Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Kommunikáció

2
Sejtek közötti kommunikáció
2/15 Sejtek közötti kommunikáció soksejtűekben elengedhetetlen összehangolni a sejtek működését direkt és indirekt kommunikáció direkt kommunikáció: rés-illeszkedés (gap junction) 6 connexin = 1 connexon; 2 connexon = 1 pórus pórusátmérő 1,5 nm, kis szerves molekulák (1500 Ms) (IP3, cAMP, peptidek) is átjutnak ingerlékeny sejteknél elektromos szinapszis a neve (gerinctelenek, szívizom, simaizom, stb.) gyors és biztos átvitel - menekülő reakcióknál gyakori: Aplysia tintalövelő sejtek, Mauthner sejt inputja, rák potroh csapás, stb. az összekapcsolt sejtek ingerküszöbe magas Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig
 6 connexin = 1 connexon; 2 connexon = 1 pórus pórusátmérő 1,5 nm, kis szerves molekulák (1500 Ms) (IP3, cAMP, peptidek) is átjutnak. ingerlékeny sejteknél elektromos szinapszis a neve (gerinctelenek, szívizom, simaizom, stb.) gyors és biztos átvitel - menekülő reakcióknál gyakori: Aplysia tintalövelő sejtek, Mauthner sejt inputja, rák potroh csapás, stb. az összekapcsolt sejtek ingerküszöbe magas. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig")
3
Indirekt kommunikáció
3/15 Indirekt kommunikáció kémiai anyag - jel - közvetítésével történik jeladó - jel - csatorna - jelfogó vannak specializált jeladók (ideg-, mirigysejt), de más sejteknél is gyakori a jelkibocsátás (pl. fehérvérsejt) a jel kémiai természete sokféle lehet: biogén aminok: katekolaminok (NA, Adr, DA), indolaminok (5-HT), imidazol (hisztamin), észterek (ACh), stb. aminosavak: glu, asp, tiroxin, GABA, glicin, stb. kis peptidek, fehérjék nukleotidok és származékaik: ATP, adenozin, stb. szteránvázas vegyületek: nemi hormonok, mellékvesekéreg hormonok, stb.
, de más sejteknél is gyakori a jelkibocsátás (pl. fehérvérsejt) a jel kémiai természete sokféle lehet: biogén aminok: katekolaminok (NA, Adr, DA), indolaminok (5-HT), imidazol (hisztamin), észterek (ACh), stb. aminosavak: glu, asp, tiroxin, GABA, glicin, stb. kis peptidek, fehérjék. nukleotidok és származékaik: ATP, adenozin, stb. szteránvázas vegyületek: nemi hormonok, mellékvesekéreg hormonok, stb.")
4
Csatorna szerinti osztályozás
4/15 a csatorna szerinti a legalapvetőbb osztályozás neurokrin a jeladó idegsejt a csatorna a szinaptikus rés nm csak a posztszinaptikus sejthez jut el (fülbesúgás) a jel mediátor, vagy neurotranszmitter parakrin (autokrin) a jeladó sokféle sejt lehet a csatorna a szövetközti tér közelben lévő sejtekhez jut el (beszéd társaságban) a jelet nevezik szöveti hormonnak is endokrin a jeladó mirigysejt, vagy idegsejt (neuroendokrin) a csatorna a véráram a szervezet minden sejtjéhez eljut (rádió, TV) a jelet hormonnak nevezik Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 8-1.
 a jel mediátor, vagy neurotranszmitter. parakrin (autokrin) a jeladó sokféle sejt lehet. a csatorna a szövetközti tér. közelben lévő sejtekhez jut el (beszéd társaságban) a jelet nevezik szöveti hormonnak is. endokrin. a jeladó mirigysejt, vagy idegsejt (neuroendokrin) a csatorna a véráram. a szervezet minden sejtjéhez eljut (rádió, TV) a jelet hormonnak nevezik Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig")
5
A jelfogók típusai hidrofil jelek esetén a jelfogó a membránban
5/15 A jelfogók típusai hidrofil jelek esetén a jelfogó a membránban hidrofóboknál a plazmában előbbi meglévő fehérjék működésére hat, utóbbi új fehérjék szintézisét befolyásolja a membránreceptor internalizálódhat és lehet plazmareceptora is membránreceptorok: ioncsatorna receptor (ligandfüggő csatornák) ideg-, és izomsejteken - gyors neurotranszmisszió ionotróp receptoroknak is nevezik G-fehérjéhez kapcsolódó receptor - legelterjedtebb idegsejteken metabotróp receptornak nevezik lassabb hatás - effektor fehérje, másodlagos hírvivő katalitikus, pl. tirozin kináz - növekedési faktorok használják - foszforilációt okoz tirozin aminosavon Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig
 ideg-, és izomsejteken - gyors neurotranszmisszió ionotróp receptoroknak is nevezik. G-fehérjéhez kapcsolódó receptor - legelterjedtebb idegsejteken metabotróp receptornak nevezik lassabb hatás - effektor fehérje, másodlagos hírvivő katalitikus, pl. tirozin kináz - növekedési faktorok használják - foszforilációt okoz tirozin aminosavon. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig")
6
Neurokrin kommunikáció I.
6/15 Neurokrin kommunikáció I. Otto Loewi Vagus-Stoff békaszív + vagus ideg preparátum - ingerlés a szivet lassítja, az oldat egy másik szívhez adva szintén lassít kémiai neurotranszmisszió első demonstrációja később: ideg-izom áttevődés - hiedelem: ACh az általános serkentő transzmitter, ionotróp, Na+-K+ vegyes csatornán át (< 1 ms) később: gátló transzmitterek Cl- csatornán át még később: lassú transzmisszió (több 100 ms), G-fehérje mechanizmussal neuromodulátor ha parakrin módon is hat egy idegvégződés egy transzmittert szabadít fel, és az egy adott hatással rendelkezik - Dale-elv ma: kismolekula + peptid együtt lehetséges
 később: gátló transzmitterek Cl- csatornán át. még később: lassú transzmisszió (több 100 ms), G-fehérje mechanizmussal. neuromodulátor ha parakrin módon is hat. egy idegvégződés egy transzmittert szabadít fel, és az egy adott hatással rendelkezik - Dale-elv. ma: kismolekula + peptid együtt lehetséges.")
7
Neurokrin kommunikáció II.
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig 7/15 Neurokrin kommunikáció II. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-31,34. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig gyors szinapszis - például az izom véglemez , kurare - dél-amerikai nyílméreg hatóanyaga mint kutatási eszköz ált. agonisták és antagonisták igen fontosak megfordulási v. reverzál potenciál - milyen ion EPSP = excitatory synaptic potential IPSP = inhibitory synaptic potential hatás a gradienstől függ - pl. Cl- Cl- csatorna nyitása akkor is lehet gátló, ha nem okoz hiperpolarizációt - membrán söntölés preszinaptikus vs. posztszinaptikus gátlás a transzmitter felszabadulása kvantált: Katz (1952) - miniatür véglemez potenciálok - Ca++ hiány esetén, ingerlésre is ennek n-szerese kb ACh molekula - 1 vezikulum hatás eliminációja
 - miniatür véglemez potenciálok - Ca++ hiány esetén, ingerlésre is ennek n-szerese. kb ACh molekula - 1 vezikulum. hatás eliminációja ")
8
Az idegsejt integratív funkciója
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig 8/15 Az idegsejt integratív funkciója Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-1. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig a jeltovábbítás során gradált és minden-vagy-semmi elektromos jelek és kémiai jelek váltogatják egymást a KIR-ben az idegsejt integrálja a hatásokat térbeli szummáció, lásd térállandó előjel, távolság az eredési dombtól fontos szempont időbeli szummáció, lásd időállandó az eredő potenciál nagysága frekvencia-kóddal továbbítva - időbeli szummációt okozhat kolokalizált transzmitterek leadása - bonyolult kölcsönhatás lehet Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-40,41. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

9
Plasztikusság a szinapszisban
9/15 Plasztikusság a szinapszisban a tanulás, a memória alapja a neuronális plasztikusság plasztikusság kell a speciális mozgások (pl. borotválkozás, tenisz, stb.) tanulásához szokások kialakulása is plasztikusságon alapul egyedfejlődés során is szükség van rá, egyes kapcsolatok megszűnnek alapja mindig a posztszinaptikus sejtből jövő visszajelzés kifejlett élőlényben: szinaptikus hatékonyság változása
 tanulásához. szokások kialakulása is plasztikusságon alapul. egyedfejlődés során is szükség van rá, egyes kapcsolatok megszűnnek. alapja mindig a posztszinaptikus sejtből jövő visszajelzés. kifejlett élőlényben: szinaptikus hatékonyság változása.")
10
D.O. Hebb posztulátuma (1949)
10/15 D.O. Hebb posztulátuma (1949) minden izgalmi szinapszis hatékonysága nő, ha a szinapszis aktivitása konzisztens, és pozitív módon korrelál a posztszinaptikus sejt aktivitásával
 minden izgalmi szinapszis hatékonysága nő, ha a szinapszis aktivitása konzisztens, és pozitív módon korrelál a posztszinaptikus sejt aktivitásával.")
11
A hatékonyságnövekedés módjai
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig.6-48. 11/15 A hatékonyságnövekedés módjai Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig preszinaptikus és posztszinaptikus mechanizmus lehetséges utóbbira nézve kevés információ van homoszinaptikus moduláció homoszinaptikus facilitáció béka izom, kuráre AP gátlására gyors ingerpár hatására ms hosszú, térbeli szummációt meghaladó növekedés a második EPSP-ben mechanizmusa a Ca++ növekedés a preszinapszisban poszttetanikus potenciáció béka izom inger sorozattal ingerelve depresszió, majd percekig tartó facilitáció mechanizmusa: a hólyagocskák mind kiürülnek (depresszió), majd helyreállítódnak, miközben a Ca++ koncentráció még magas Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig
, majd helyreállítódnak, miközben a Ca++ koncentráció még magas. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig")
12
Heteroszinaptikus moduláció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig 12/15 Heteroszinaptikus moduláció a transzmitter felszabadulást más szinapszis vagy a keringés által odajuttatott modulátor befolyásolja pl. szerotonin - puhatestűek és gerincesek oktopamin - rovarok NA és GABA - gerincesek idetartozik a preszinaptikus gátlás is serkentő hatású modulációk heteroszinaptikus facilitáció - Aplysia - érző és motoros idegsejt közötti áttevődés fokozódik 5-HT jelenlétében mechanizmus: 5-HT - cAMP - KS-csatorna záródik - AP hosszabb, több Ca++ lép be long-term potenciáció - LTP pl. hippokampusz intenzív ingerlésre órákig, napokig, hetekig tartó hatásnövekedés mindig NMDA szerepel Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

13
G-fehérjéhez kapcsolt hatás
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 9-8. 13/15 G-fehérjéhez kapcsolt hatás Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 9-9b. ismétlés: jel szerinti osztályozás lipofil jel (szteroid, tiroxin) - plazmareceptor - fehérjeszintézis módosítás lipofób jel (a többi) - membránreceptor - meglévő fehérje módosítása ioncsatorna (ligand-függő) - ionotróp receptor G-fehérjéhez kapcsolódó 7M receptor - metabotróp receptor katalitikus receptor - tirozin kináz, cGMP szintetáz a legelterjedtebb a G-fehérjéhez kötött ligand + receptor = aktivált receptor aktivált receptor + G-fehérje = aktivált G-fehérje (GDP - GTP csere) aktivált G-fehérje - -alegység disszociál -alegység - effektor fehérje aktiváció -alegység - GTP lebontás GDP-vé - hatás vége
 - plazmareceptor - fehérjeszintézis módosítás lipofób jel (a többi) - membránreceptor - meglévő fehérje módosítása. ioncsatorna (ligand-függő) - ionotróp receptor. G-fehérjéhez kapcsolódó 7M receptor - metabotróp receptor. katalitikus receptor - tirozin kináz, cGMP szintetáz. a legelterjedtebb a G-fehérjéhez kötött. ligand + receptor = aktivált receptor. aktivált receptor + G-fehérje = aktivált G-fehérje (GDP - GTP csere) aktivált G-fehérje - -alegység disszociál. -alegység - effektor fehérje aktiváció. -alegység - GTP lebontás GDP-vé - hatás vége.")
14
Effektor fehérjék K+-csatorna - nyitás, vagy csukás
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig Alberts et al.: Molecular biology of the cell, Garland Inc., N.Y., London 1989, Fig 14/15 Effektor fehérjék Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 1-4. Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig K+-csatorna - nyitás, vagy csukás másodlagos hírvivőn keresztüli hatás Sutherland Nobel-díj - cAMP rendszer további másodlagos hírvivők felfedezése hatásmechanizmusok: cAMP IP3 - diacilglicerol Ca++ egy jel, több lehetséges útvonal egy útvonal, több lehetséges jel jelentőség: jel felerősítése a receptor jelenléte, és milyensége a döntő: pl. szerotonin receptorok Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

15
Katalitikus receptorok
15/15 Katalitikus receptorok Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

17
Gap junction (rés-illeszkedés)
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

18
Csatorna szerinti felosztás
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 8-1.

19
Gyors és lassú neurotranszmisszió
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

20
A neuromuszkuláris junkció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

21
A neuromuszkuláris véglemez
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

22
A jel eliminációja Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-31,34.

23
Az ingerület terjedése KIR-ben
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-1.

24
AP generálása az axondombon
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

25
Térbeli szummáció Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

26
EPSP és IPSP szummációja
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

27
Időbeli szummáció Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

28
Frekvenciakód Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

29
Neuromoduláció Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 6-40,41.

30
Homoszinaptikus facilitáció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig.6-48.

31
A facilitáció Ca++ függése
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

32
Poszttetanikus potenciáció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

33
Heteroszinaptikus facilitáció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

34
Long-term potenciáció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

35
Hatásmód és zsíroldékonyság
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 9-8.

36
Zsíroldékony jelek hatásmódja
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 9-9b.

37
Effektor fehérje: K+-csatorna
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

38
Másodlagos hírvivők Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

39
cAMP szignalizáció Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

40
Inozitoltrifoszfát szignalizáció
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

41
Ca++ szignalizáció Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig

42
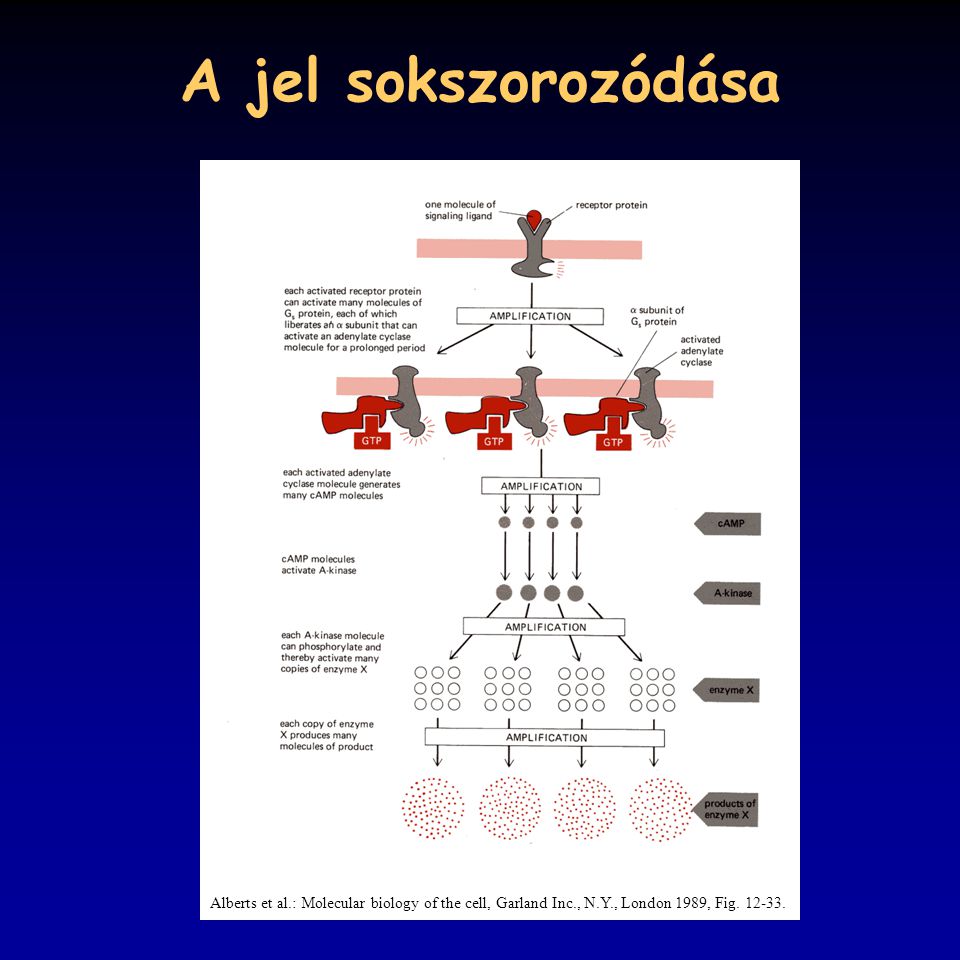
A jel sokszorozódása Alberts et al.: Molecular biology of the cell, Garland Inc., N.Y., London 1989, Fig
43
Szerotonin receptorok
Eckert: Animal Physiology, W.H.Freeman and Co., N.Y.,2000, Fig. 1-4.

Hasonló előadás

















>")
>")

