Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Forrongó evolúcióbiológia
Fajképződés ökológus szemmel Pásztor Erzsébet ELTE

2
Paradigmaváltás van a fajképződés kutatásában.
Üzenet Paradigmaváltás van a fajképződés kutatásában. Genetikai Ökológiai paradigma

3
Elméletek genetikai ökológiai Szintetikus evolúcióelmélet
szaporodási izoláció (biológiai fajfogalom) allopatrikus fajképződés Adaptív speciáció elmélete gyakoriságfüggő szelekció (szétválasztó) szimpatrikus fajképződés genetikai ökológiai
 allopatrikus fajképződés. Adaptív speciáció elmélete. gyakoriságfüggő szelekció (szétválasztó) szimpatrikus fajképződés. genetikai. ökológiai.")
4
A biológiai fajfogalom bajban van!
Genom elemzések eredményei „fajok” közti hibridizációk nyomai

5
Elméletek genetikai ökológiai Szintetikus evolúcióelmélet
Hogyan alakulnak ki szaporodásilag izolált csoportok? Ökológiai izoláció, ökológiai speciáció - Adaptív speciáció elmélete Hogyan hoz létre a természetes szelekció evolúciós elágazásokat? genetikai ökológiai

6
Mérföldkövek Mayr, E. 1963: Animal species and evolution
Otte, Endler 1989: Speciation and its consequences Coyne, Orr 2003: Speciation. Howard, Berlocher 1998: Endless forms. Species and speciation Schluter, D. 2000: The ecology of adaptive radiation Dieckmann et. al 2004: Adaptive speciation

7
Amiről szó lesz Régi-új fajképződés elmélet Új evidenciák Ökológiai újdonságok

8
Az adaptív fajképződés fogalma
Darwin is megmondta... Az egymástól leginkább különböző, különböző növekedési korlátba ütköző változatok tudják számukat egymás jelenlétében növelni, azaz fennmaradni. Érv: egy terület annál több élőlényt tud eltartani, mennél különbözőbbek,

9
Az adaptív fajképződés fogalma
„Gyakoriságfüggő biológiai kölcsönhatásokhoz kapcsolódó szétválasztó szelekció következtében létrejövő fajképződés” Dieckmann et. al 2004

10
Gyakoriságfüggő szelekció
Az egyes változatok rátermettsége a populáció összetételének a függvénye. Az egyes allélok átlagos rátermettsége (marginális rátermettség) a relatív gyakoriságuk függvénye. Tétel: Gyakoriságfüggő szelekció esetén, egyensúlyi állapotban minden allél rátermettsége azonos.
 a relatív gyakoriságuk függvénye. Tétel: Gyakoriságfüggő szelekció esetén, egyensúlyi állapotban. minden allél rátermettsége azonos.")
11
Drosophila melanogaster Lárva táplálkozási gén: forR :”görgő”
Maintaining a behaviour polymorphism by frequency-dependent selection on a single gene Nature 447, (10 May 2007) Drosophila melanogaster Lárva táplálkozási gén: forR :”görgő” forS :”ülő” cGMP függő protein-kináz szintje más mozgásmennyiség, forma más lárva stádiumban versengés van a táplálékért
 Drosophila melanogaster. Lárva táplálkozási gén: forR : görgő forS : ülő cGMP függő protein-kináz szintje más. mozgásmennyiség, forma más. lárva stádiumban versengés van a táplálékért.")
12
Csak a két homozigóta típus szerepel!
Maintaining a behaviour polymorphism by frequency-dependent selection on a single gene Nature 447, (10 May 2007) Kísérlet: faktorok Lárva denzitás/forrásmennyiség (20 ill. 40 db) Relatív gyakoriságok változtatása Csak a két homozigóta típus szerepel! (III. Kromoszómán lokalizált zöld fluorescens fehérjével jelölték a homozigótákat) Rátermettség mértéke: bebábozódó lárvák aránya 1 ismétlés
 Kísérlet: faktorok. Lárva denzitás/forrásmennyiség (20 ill. 40 db) Relatív gyakoriságok változtatása. Csak a két homozigóta típus szerepel! (III. Kromoszómán lokalizált zöld fluorescens fehérjével jelölték a homozigótákat) Rátermettség mértéke: bebábozódó lárvák aránya. 1 ismétlés.")
13
frequency x genotype interaction F2,2 = 96.789, p = 0.010!
görgő ülő alacsony denzitás görgő ülő magas denzitás

14
Nem kapcsoltság! forR genomban forS mutáns ugyanígy viselkedik!
jelölt, jelöletlen ugyanígy viselkedik!

15
Az adaptív tájkép metafora
Optimalizáció Evolúció=hegymászás út=idő

16
Szétválasztó szelekció
rátermettség tulajdonság

17
Az adaptív tájkép metafora
hágó

18
De másból azért meg lehet gazdagodni!
Az aranyláz metafora Versengés az aranyért = versengés a forrásokért A túl sok aranyásó, elhordja a hegyet! De másból azért meg lehet gazdagodni!

19
Az adaptív tájkép állandóan változik!
Jobb metafora?

20
Szaporodási izoláció? Szétágazás ökológiai lehetősége?
A fajképződés motorja Szaporodási izoláció? Szétágazás ökológiai lehetősége?

21
A fajképződés gyakorisága függ az ökológiai körülményektől
Magasabb kolonizáció távoli földrészekre, szigetekre, tavakba – interspecifikus kompetíció, predáció, betegségek csökkent mértékűek tömeges kihalások után korábban kihasználatlan forrásokhoz való adaptáció után új források tömegének megjelenése után

22
A növekedés-elágazás metafora
növekedés = egyirányú szelekció elágazás = fajképződés A szigeteken nincsenek versenytárs fajok! Ha sok a versenytárs, nincs elágazás!

23
A fajképződés folyamata
Allopatrikus Szimpatrikus Szaporodási izoláció Ökológiai differenciálódás - szétválasztó szelekció Ökológiai különbségek Izoláció megerősítése Szelekció szaporodási izolációra Szétterjedés 1. 2. 3.

24
Szintetikus evolúcióelmélet (Ernst Mayr)
szaporodási izoláció a folyamat elején elengedhetetlen a fajképződéshez, mert az érintett lokuszokon lévő allélok rekombinációja miatt egyébként nehezen indul be földrajzi izoláció elsődleges jelentősége: szaporodási egységet biztosító mechanizmusok erodálása, megváltozása

25
A helyzet Szimpatrikus speciációt Mayr és követői szinte lehetetlennek tartották. Mayr halála óta gyorsuló ütemben halmozódnak a példák szimpatrikus illetve parapatrikus speciációra.

26
Anolis gyíkok esete Kis Antillák
2-5 millió éves szigetek ~ egy sziget - egy faj fajon belül hegyi és parti ökotípus Tipikus allopatrikus fajképződés. Tipikus parapatrikus fajképződés.

27
Szaporodási izolációval induló fajképződés

28
Szaporodási izoláció Géncsere megakadályozása a különböző típusok között prezigótikus fizikai gát időbeli elkülönülés: pl. virágzás időzítése térbeli elkülönülés egy élőhelyen viselkedési mechanizmusra épülő asszortatív párosodás posztzigótikus életképtelen hibrid: egyedfejlődés különböző stádiumokban akadhat el steril hibrid

29
Prezigotikus izoláció egy lépésben
Rei Ueshima*†, Takahiro Asami†‡ NATURE|VOL 425 | 16 OCTOBER 2003 |

30
Egy lokuszos fajképződés!
Egyetlen dexter Bradybaena similaris, minden utódja sinisterheterozigóták=F1 F2: 101 dexter 30 sinistral (H0= 3:1 χ2 = 3.01, p=0.62 Genetika gyakorlati jegyzet, Drosophila fejezet). dexter domináns allél
. dexter domináns allél.")
31
Erős szelekció dexter formára
dexter ősi forma 4 sinistral faj monofiletikus, azaz egyetlen mutációra vezethető vissza! Visszamutáció+fajképződés 3 esetben történt A domináns dexter mutáns allél könnyebben fixálódik, mint recesszív párja gyakoriságfüggés miatt

32
Mágikus tulajdonságokon alapuló fajképződés
Mágikus tulajdonság az, ami egyszerre vezet pozitív asszortatív párosodáshoz (hasonló hasonlót választ) és niche szegregációhoz.
 és niche szegregációhoz.")
33
Pre-zigotikus izoláció és táplálék divergencia együttes kialakulása
Patkósorrú denevér család (Rhinolophidae) 5 millió év, 30 faj Rhinolophus philippinensis 3 szimpatrikus forma: kicsi, közepes, nagy
 5 millió év, 30 faj. Rhinolophus philippinensis. 3 szimpatrikus forma: kicsi, közepes, nagy.")
34
Harmonic-hopping in Wallacea’s bats Tigga Kingston & Stephen J
Harmonic-hopping in Wallacea’s bats Tigga Kingston & Stephen J. Rossiter Különböző frekvenciájú hangot adó denevérek tápláléka különbözik: Nagy denevér – nagy táplálék! Különböző frekvenciájú hangot adó denevérek nem hallják egymást! Genetikai izoláció kimutatható microsatellit lokusz alapján! NATURE | VOL 429 | 10 JUNE 2004 |

35
Pre-zigotikus izoláció és táplálék divergencia együttes kialakulása
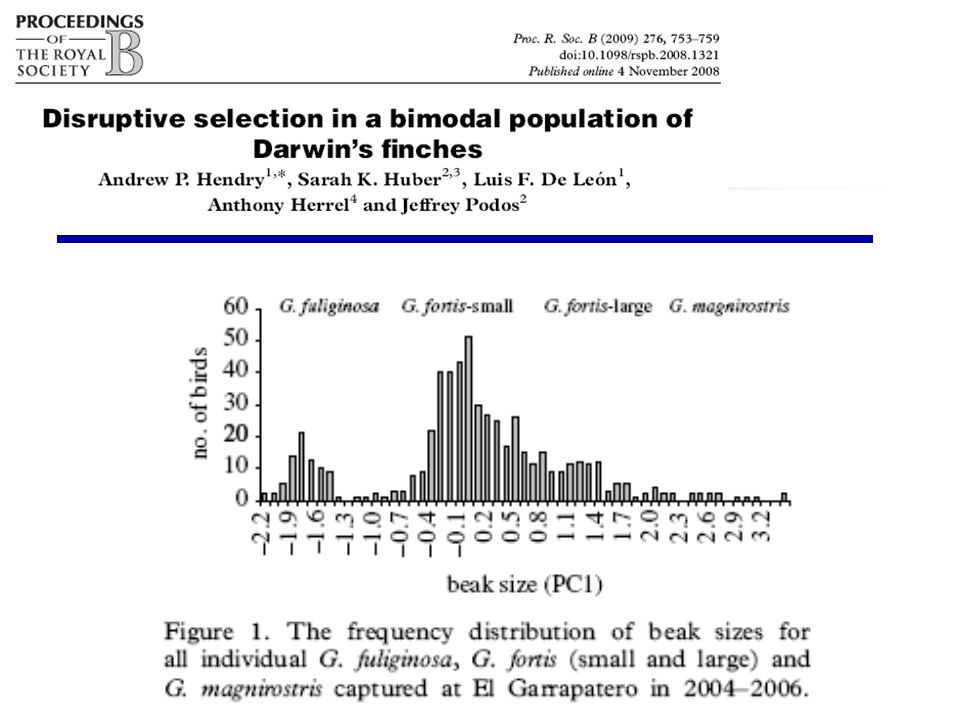
Geospiza fortis magevő darwin pinty Csőrméret magméret ének táplálék izoláció szap izoláció

36
A csőr alakjára ható molekuláris mechanizmusok

37
Katasztrofális időjárás okozta irányító szelekció
Katasztrofális időjárás okozta irányító szelekció. Darwin pintyek, aszály, csőr és testméret nagyobbodás

39
Szétválasztó szelekció kimutatása
Csőrméret, Galapagos, közép földi pinty

40
Méret szerinti asszortatív párosodás

41
A fenotípusok rátermettsége gyakoriságfüggő: Dactylorhiza sambucina (bodzaszagú ujjaskosbor) virágszín polimorfizmusának esete A poszméhek érzékelik a visszavert fény arányát. A kétféle színű virág fényvissza- verése más hullámhossz eloszlást mutat. Negative frequency-dependent selection maintains a dramatic flower color polymorphism in the rewardless orchid Dactylorhiza sambucina (L.) Soò in:PNAS 2001
 Soò in:PNAS")
42
A cikk ajánlott olvasmány!
A fenotípusok rátermettsége gyakoriságfüggő: Dactylorhiza sambucina (bodzaszagú ujjaskosbor) virágszín polimorfizmusának esete Nem ad nektárt, naív poszméhek a ritkább változatokat gyakrabban látogatják. Az adatokból becsülhető egyensúlyi gyakoriságokat szaggatott vonal jelöli. A cikk ajánlott olvasmány! Pollinium: Relative male (a) and female (b) reproductive success of the yellow morph as a function of the relative frequency of the yellow morph in each array. Male reproductive success for a morph was quantified as the average proportion of pollinia removed from plants within each array (n = 5 to 45 plants) (●). Female reproductive success was measured as the average proportion of stigmas receiving pollen ( ) and setting fruits (□) for plants within each array. Relative reproductive success was calculated as described in Materials and Methods. Each data point represents the mean of two arrays per frequency, with bars showing one SEM. Statistics were calculated by using all values. r is the Spearman correlation coefficient (Pearson correlation coefficients were identical). *, P < 0.05; **, P < The slopes given are the lines fitted in regression analysis: (a) y = −0.66x ; (b1, solid line for pollinia deposition) y = −0.63x , and (b2, broken line for fruit set) y = −0.59x The horizontal line corresponds to equal reproductive success between the two morphs. The intersection between regression lines and the horizontal line gives the value of predicted morph frequencies at equilibrium (represented by vertical dotted lines). We constructed 10 synthetic arrays of D. sambucina, varying the morph frequency within the arrays to test the influence of morph frequency on relative reproductive success. The experiment was performed in France during May and June 2000 on the central part of the Causse Noir (N 44° 10′, E 03° 20′, Altitude: 1,000 ± 11 m) in the Massif des Cévennes where D. sambucina does not occur naturally, although populations are recorded frequently elsewhere in the Massif and at the edge of the Causse (minimum 5 km from the experimental areas). We chose such isolation to ensure naivete of the pollinators to D. sambucina. The predominant pollinators of D. sambucina are bumblebees, principally Bombus lapidarius (6) and Bombus terrestris (L.G. and A.S., personal observation), which were abundant in the experimental areas. We removed 600 individual plants from a population ≈12 km from the Causse and potted them. Experimental arrays then were constructed in 10 locations on the Causse. Each location was chosen for habitat similarity both to each other and to that of natural populations. A minimum distance of 2 km separated the locations to ensure isolation of the pollinator fauna. Arrays consisted of 50 plants dispersed on a square grid. Grid size was 9.5 by 9.5 m, with grid spacing of 0.5 m, giving 400 potential positions for plants. Plants were allocated randomly to arrays after we controlled for flower number, reallocating plants among arrays where necessary. Plants then were allocated randomly to the 400 potential positions within arrays. Two replicates each of 0.1, 0.3, 0.5, 0.7, and 0.9 yellow morphs were used, with treatments randomly allocated to arrays. We commenced experiments on May 6th, 2000, the peak flowering time for D. sambucina in surrounding natural plant populations. After 5 days, the number of pollinia removed and the number of stigmas with pollen depositions were quantified (all pollinator visitation had occurred by this point). Six weeks later, fruit set was assessed by counting the total number of fruits produced per plant.
 virágszín polimorfizmusának esete. Nem ad nektárt, naív poszméhek a ritkább változatokat gyakrabban látogatják. Az adatokból becsülhető egyensúlyi gyakoriságokat szaggatott vonal jelöli. A cikk ajánlott olvasmány! Pollinium: Relative male (a) and female (b) reproductive success of the yellow morph as a function of the relative frequency of the yellow morph in each array. Male reproductive success for a morph was quantified as the average proportion of pollinia removed from plants within each array (n = 5 to 45 plants) (●). Female reproductive success was measured as the average proportion of stigmas receiving pollen ( ) and setting fruits (□) for plants within each array. Relative reproductive success was calculated as described in Materials and Methods. Each data point represents the mean of two arrays per frequency, with bars showing one SEM. Statistics were calculated by using all values. r is the Spearman correlation coefficient (Pearson correlation coefficients were identical). *, P < 0.05; **, P < The slopes given are the lines fitted in regression analysis: (a) y = −0.66x ; (b1, solid line for pollinia deposition) y = −0.63x , and (b2, broken line for fruit set) y = −0.59x The horizontal line corresponds to equal reproductive success between the two morphs. The intersection between regression lines and the horizontal line gives the value of predicted morph frequencies at equilibrium (represented by vertical dotted lines). We constructed 10 synthetic arrays of D. sambucina, varying the morph frequency within the arrays to test the influence of morph frequency on relative reproductive success. The experiment was performed in France during May and June 2000 on the central part of the Causse Noir (N 44° 10′, E 03° 20′, Altitude: 1,000 ± 11 m) in the Massif des Cévennes where D. sambucina does not occur naturally, although populations are recorded frequently elsewhere in the Massif and at the edge of the Causse (minimum 5 km from the experimental areas). We chose such isolation to ensure naivete of the pollinators to D. sambucina. The predominant pollinators of D. sambucina are bumblebees, principally Bombus lapidarius (6) and Bombus terrestris (L.G. and A.S., personal observation), which were abundant in the experimental areas. We removed 600 individual plants from a population ≈12 km from the Causse and potted them. Experimental arrays then were constructed in 10 locations on the Causse. Each location was chosen for habitat similarity both to each other and to that of natural populations. A minimum distance of 2 km separated the locations to ensure isolation of the pollinator fauna. Arrays consisted of 50 plants dispersed on a square grid. Grid size was 9.5 by 9.5 m, with grid spacing of 0.5 m, giving 400 potential positions for plants. Plants were allocated randomly to arrays after we controlled for flower number, reallocating plants among arrays where necessary. Plants then were allocated randomly to the 400 potential positions within arrays. Two replicates each of 0.1, 0.3, 0.5, 0.7, and 0.9 yellow morphs were used, with treatments randomly allocated to arrays. We commenced experiments on May 6th, 2000, the peak flowering time for D. sambucina in surrounding natural plant populations. After 5 days, the number of pollinia removed and the number of stigmas with pollen depositions were quantified (all pollinator visitation had occurred by this point). Six weeks later, fruit set was assessed by counting the total number of fruits produced per plant.")
43
Fajképződés térbeli elkülönüléssel –
parapatrikus fajképződés Fajképződés térbeli elkülönüléssel –

44
Amphilophus citrinellus, általánosan elterjedt: bentikus
Sympatric speciation in Nicaraguan crater lake cichlid fish Marta Barluenga, Kai N. Stölting, Walter Salzburger, Moritz Muschick & Axel Meyer Amphilophus citrinellus, általánosan elterjedt: bentikus Amphilophus zaliosus egyetlen izolált kráter tóban: hosszabb testű, limnetikus mtDNS, mikroszatellita, RFLP, ökomorfológia Környező hegyek tavainak ciklidái Nature Vol 439|9 February 2006|

45
Sympatric speciation in Nicaraguan crater lake cichlid fish Marta Barluenga, Kai N. Stölting, Walter Salzburger, Moritz Muschick & Axel Meyer

47
Különbség a testfelépítésben

48
Különbség az állkapocs alakjában

49
Különbség a gyomor tartalomban

50
Ciklidák sekélyvizű afrikai tavakban
Szaporodási izoláció vizuális jegyeken alapuló párválasztás alapján Ökológiai differenciáció 500 faj max év Viktória tó Hogyan jött létre a környezettel változó rátermettségű vizuális jegyeken alapuló párválasztás?

51
Lord Howe Island Howea forsteriana H. belmoreana

52
Sympatric speciation in palms on an oceanic island Vincent Savolainen, Marie-Charlotte Anstett, Christian Lexer, Ian Hutton, James J. Clarkson, Maria V. Norup, Martyn P. Powell, David Springate, Nicolas Salamin & William J. Baker Különbség a virágzás időzítésében: szaporodási izoláció Nature Vol 441|11 May 2006|

53
Niche különbség (szegregáció)
")
54
Genetikai különbségek

55
A két kovamoszat faj funkcionális válasza a szilikát koncentrációra

56
A forráskoncentráció csökken és stabilizálódik két kovamoszat faj versengése közben
Az a változat nyer, amelyiknek magasabb a növekedési rátája alacsony szilikát koncentráción.

57
Hőmérséklet tolerancia és az R* hőmérsékletfüggése
R* hőmérsékletfüggése a két kovamoszat fajban Hőmérséklet tolerancia a két kovamoszat fajban

Hasonló előadás



 pintyek>")










 Bihari Péter.>")





