Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Sejten belüli jelátadás – jeltovábbitás a citoplazmában

2
Sejten belüli másodlagos hírvivő molekulák
Structural formulas of four common intracellular second messengers. Their abbreviations are indicated. The calcium ion (Ca2+) and several membrane-bound inositol phospholipids (phosphoinositides) also act as second messengers but are not shown Ca2+ ionok, sejtmembránhoz kötött inozitol foszfolipidek (foszfoinozitidek)
 and several membrane-bound inositol phospholipids (phosphoinositides) also act as second messengers but are not shown. Ca2+ ionok, sejtmembránhoz kötött inozitol foszfolipidek (foszfoinozitidek)")
3
Általános, sejten belüli jelátadó molekulák
GTP kötő fehérjék: molekuláris kapcsolók Adapter fehérjék Trimer (a,b,g) Monomer (ras, rho) Ras = rat sarcoma (retrovírus indukált) Protein kinázok Multiprotein szignál komplexek Common intracellular signaling proteins. (a) GTP-binding proteins with GTPase activity function as molecular switches. When bound to GTP they are active; when bound to GDP, they are inactive. They fall into two categories, trimeric G proteins and Ras-like proteins. (b) Protein kinases modulate the activity or the binding properties of substrate proteins by phosphorylating serine, threonine, or tyrosine residues. The phosphorylated form of some proteins is active, whereas the dephosphorylated form of other proteins is active. The combined action of kinases and phosphatases, which dephosphorylate specific substrates, can cycle proteins between active and inactive states. (c) Adapter proteins contain various protein-binding motifs that promote the formation of multiprotein signaling complexes. Aktív vagy inaktív Aktív vagy inaktív Protein tirozin kinázok Szerin/treonin kinázok
 Monomer (ras, rho) Ras = rat sarcoma (retrovírus indukált) Protein kinázok. Multiprotein szignál komplexek. Common intracellular signaling proteins. (a) GTP-binding proteins with GTPase activity function as molecular switches. When bound to GTP they are active; when bound to GDP, they are inactive. They fall into two categories, trimeric G proteins and Ras-like proteins. (b) Protein kinases modulate the activity or the binding properties of substrate proteins by phosphorylating serine, threonine, or tyrosine residues. The phosphorylated form of some proteins is active, whereas the dephosphorylated form of other proteins is active. The combined action of kinases and phosphatases, which dephosphorylate specific substrates, can cycle proteins between active and inactive states. (c) Adapter proteins contain various protein-binding motifs that promote the formation of multiprotein signaling complexes. Aktív vagy inaktív. Aktív vagy inaktív. Protein tirozin kinázok. Szerin/treonin kinázok.")
4
G fehérjékhez kötött receptorok és receptor tirozin kinázok jelátviteli sémájának összehasonlítása
Protein kinázokhoz kapcsolt receptorok: BCR, TCR, citokin R kemokin receptorok Schematic overview of common signaling pathways downstream from G protein – coupled receptors (GPCRs) and receptor tyrosine kinases (RTKs). Hormone binding to the receptor initiates a series of events leading to phosphorylation of specific substrate proteins, which mediate the cellular responses such as changes in the activity of metabolic enzymes, gene expression, and cytoskeletal structures. The kinase cascade entails sequential activation of specific protein kinases induced by a signal from activated Ras protein. Second messengers (SM) play a role in some RTK signaling pathways, although not in the pathway depicted here. Likewise, some GPCR pathways do not involve second messengers; these lead to activation of MAP kinase. Metabolikus enzimek aktivitása Gén expresszió Citoszkeletális struktúrák
 and receptor tyrosine kinases (RTKs). Hormone binding to the receptor initiates a series of events leading to phosphorylation of specific substrate proteins, which mediate the cellular responses such as changes in the activity of metabolic enzymes, gene expression, and cytoskeletal structures. The kinase cascade entails sequential activation of specific protein kinases induced by a signal from activated Ras protein. Second messengers (SM) play a role in some RTK signaling pathways, although not in the pathway depicted here. Likewise, some GPCR pathways do not involve second messengers; these lead to activation of MAP kinase. Metabolikus enzimek aktivitása. Gén expresszió. Citoszkeletális struktúrák.")
5
G-fehérjéhez kapcsolodó receptorok jelátvitele az adenilcikláz aktiválódásán keresztül
Activation of adenylyl cyclase following binding of an appropriate hormone (e.g., epinephrine, glucagon) to a Gs protein – coupled receptor. Following ligand binding to the receptor, the Gs protein relays the hormone signal to the effector protein, in this case adenylyl cyclase. Gs cycles between an inactive form with bound GDP and an active form with bound GTP. Dissociation of the active form yields the Gsα · GTP complex, which directly activates adenylyl cyclase. Activation is short-lived because GTP is rapidly hydrolyzed (step 5 ). This terminates the hormone signal and leads to reassembly of the inactive Gs · GDP form, returning the system to the resting state. Binding of another hormone molecule causes repetition of the cycle. Both the Gγ and Gsα subunits are linked to the membrane by covalent attachment to lipids. Binding of the activated receptor to Gsα promotes dissociation of GDP and its replacement with GTP. gyors
 to a Gs protein – coupled receptor. Following ligand binding to the receptor, the Gs protein relays the hormone signal to the effector protein, in this case adenylyl cyclase. Gs cycles between an inactive form with bound GDP and an active form with bound GTP. Dissociation of the active form yields the Gsα · GTP complex, which directly activates adenylyl cyclase. Activation is short-lived because GTP is rapidly hydrolyzed (step 5 ). This terminates the hormone signal and leads to reassembly of the inactive Gs · GDP form, returning the system to the resting state. Binding of another hormone molecule causes repetition of the cycle. Both the Gγ and Gsα subunits are linked to the membrane by covalent attachment to lipids. Binding of the activated receptor to Gsα promotes dissociation of GDP and its replacement with GTP. gyors.")
6
Az adenilcikláz Gsα és Giα fehérjékhez kötődő receptorokon keresztüli szabályozása
Hormon-induced activation and inhibition of adenylyl cyclase is mediated by Gsα (blue) and Giα (brown), respectively. Binding of Gsα · GTP to adenylyl cyclase activates the enzyme (see Figure 20-16), whereas binding of Giα inhibits adenylyl cyclase. The Gβγ subunit in both stimulatory and inhibitory G proteins is identical; the Gα subunits and the receptors differ. Some isoforms of adenylyl cyclase are directly inhibited by binding to Gβγ. Others require coincident binding of dissociated Gsα=GTP and Gβγ subunits adenylyl cyclase adenylyl cyclase aktiválás gátlás
 and Giα (brown), respectively. Binding of Gsα · GTP to adenylyl cyclase activates the enzyme (see Figure 20-16), whereas binding of Giα inhibits adenylyl cyclase. The Gβγ subunit in both stimulatory and inhibitory G proteins is identical; the Gα subunits and the receptors differ. Some isoforms of adenylyl cyclase are directly inhibited by binding to Gβγ. Others require coincident binding of dissociated Gsα=GTP and Gβγ subunits. adenylyl cyclase adenylyl cyclase aktiválás gátlás.")
7
A trimer GTP-t kötő fehérje (G fehérje) szerkezete
b Kapcsolódás a sejtmembránhoz Nukleotid (GTP) kötőhely, A trimeric GTP-binding protein, or G protein. This type of G protein functionally couples transmembrane receptors to either enzymes or ion channels in the plasma membrane. (Based on D.G. Lombright et al., Nature 379:311–319, 1996.) g A transzmembrán receptorokat enzimekhez vagy ioncsatornákhoz kapcsolja (Based on D.G. Lombright et al., Nature 379:311–319, 1996.)
 kötőhely, A trimeric GTP-binding protein, or G protein. This type of G protein functionally couples transmembrane receptors to either enzymes or ion channels in the plasma membrane. (Based on D.G. Lombright et al., Nature 379:311–319, 1996.) g. A transzmembrán receptorokat enzimekhez vagy ioncsatornákhoz kapcsolja (Based on D.G. Lombright et al., Nature 379:311–319, 1996.)")
8
Ras-szerű domén (GTPáz aktivitás)
A GTP kötő Gs szerkezete komplexben az adenililcikláz katalitikus doménjének két fragmentjével Adenilil cikláz Ras-szerű domén (GTPáz aktivitás) The structure of Gsα · GTP complexed with two fragments encompassing the catalytic domain of adenylyl cyclase. The α3-β5 loop and the helix in the switch II region (red) of Gsα · GTP interact simultaneously with a specific region of adenylyl cyclase. The dark gray portion of Gsα is the Ras-like domain, which has GTPase activity, and the light gray portion is the helical domain (see Figure 20-19). The two adenylyl cyclase fragments are shown in purple and brown. Forskolin locks the cyclase fragments in their active conformations. Other studies indicate that Giα binds to a different region of adenylyl cyclase, accounting for its different effect on the effector. Helikális domén (Fixálja a cikláz fragmenteket az aktív konformációban kristályosításkor)
 The structure of Gsα · GTP complexed with two fragments encompassing the catalytic domain of adenylyl cyclase. The α3-β5 loop and the helix in the switch II region (red) of Gsα · GTP interact simultaneously with a specific region of adenylyl cyclase. The dark gray portion of Gsα is the Ras-like domain, which has GTPase activity, and the light gray portion is the helical domain (see Figure 20-19). The two adenylyl cyclase fragments are shown in purple and brown. Forskolin locks the cyclase fragments in their active conformations. Other studies indicate that Giα binds to a different region of adenylyl cyclase, accounting for its different effect on the effector. Helikális domén. (Fixálja a cikláz fragmenteket az aktív konformációban kristályosításkor)")
9
A ras kis G fehérje aktiválódásának 4 lépése
Cycling of the Ras protein between the inactive form with bound GDP and the active form with bound GTP occurs in four steps. By mechanisms discussed later, binding of certain growth factors to their receptors induces formation of the active Ras · GTP complex. Step 1: Guanine nucleotide – exchange factor (GEF) facilitates dissociation of GDP from Ras. Step 2: GTP then binds spontaneously, and GEF dissociates yielding the active Ras · GTP form. Steps 3and 4: Hydrolysis of the bound GTP to regenerate the inactive Ras · GDP form is accelerated a hundredfold by GTPase-activating protein (GAP). Unlike Gα, cycling of Ras thus requires two proteins, GEF and GAP; otherwise, Gα and Ras exhibit many common features. Szabályozó fehérjék: GAP (GTPáz aktiváló protein) ras GTPase aktivitását stimulálja és GEF (guanin nukleotid csere faktor) GDP disszociálódását segíti elő
 facilitates dissociation of GDP from Ras. Step 2: GTP then binds spontaneously, and GEF dissociates yielding the active Ras · GTP form. Steps 3and 4: Hydrolysis of the bound GTP to regenerate the inactive Ras · GDP form is accelerated a hundredfold by GTPase-activating protein (GAP). Unlike Gα, cycling of Ras thus requires two proteins, GEF and GAP; otherwise, Gα and Ras exhibit many common features. Szabályozó fehérjék: GAP (GTPáz aktiváló protein) ras GTPase aktivitását stimulálja és. GEF (guanin nukleotid csere faktor) GDP disszociálódását segíti elő.")
10
Ras-GDP-Sos komplex és a ras-GTP szerkezete röntgenkrisztallográfiás mérések alapján
Structures of Ras · GDP-Sos complex and Ras · GTP determined by x-ray crystallography. (a) Sos (shown as a trace diagram) binds to two switch regions of Ras · GDP, leading to a massive conformational change in Ras. In effect, Sos pries open Ras by displacing the switch I region, thereby allowing GDP to diffuse out. (b) GTP is thought to bind to Ras-Sos first through its base; subsequent binding of the GTP phosphates complete the interaction. The resulting conformational change in Ras displaces Sos and promotes interaction of Ras · GTP with its effectors (discussed later). Ras is in a slightly different orientation in parts (a) and (b). GDP and GTP are shown as small stick models in the center of Ras; the adjacent sphere is a Mg2+ ion. Sos, megváltotatja a ras konformációját (Son of sevenles, cserefaktor (GEF))
 Sos (shown as a trace diagram) binds to two switch regions of Ras · GDP, leading to a massive conformational change in Ras. In effect, Sos pries open Ras by displacing the switch I region, thereby allowing GDP to diffuse out. (b) GTP is thought to bind to Ras-Sos first through its base; subsequent binding of the GTP phosphates complete the interaction. The resulting conformational change in Ras displaces Sos and promotes interaction of Ras · GTP with its effectors (discussed later). Ras is in a slightly different orientation in parts (a) and (b). GDP and GTP are shown as small stick models in the center of Ras; the adjacent sphere is a Mg2+ ion. Sos, megváltotatja a ras konformációját. (Son of sevenles, cserefaktor (GEF))")
11
Receptor tirozin kinázok szerkezete és aktiválódása
General structure and activation of receptor tyrosine kinases (RTKs). The ligands for some RTKs, such as the receptor for EGF depicted here, are monomeric; ligand binding induces a conformational change in receptor monomers that promotes their dimerization. The ligands for other RTKs are dimeric; their binding brings two receptor monomers together directly (see Figure 20-4d). In either case, the kinase activity of each subunit of the dimeric receptor initially phosphorylates tyrosine residues near the catalytic site in the other subunit. Subsequently, tyrosine residues in other parts of the cytosolic domain are autophosphorylated. A ligandum kötődése konformációs változást indukálhat, ami elősegíti a receptor dimer kialakulását transzfoszforiláció a két receptor lánc között autofoszforiláció
. The ligands for some RTKs, such as the receptor for EGF depicted here, are monomeric; ligand binding induces a conformational change in receptor monomers that promotes their dimerization. The ligands for other RTKs are dimeric; their binding brings two receptor monomers together directly (see Figure 20-4d). In either case, the kinase activity of each subunit of the dimeric receptor initially phosphorylates tyrosine residues near the catalytic site in the other subunit. Subsequently, tyrosine residues in other parts of the cytosolic domain are autophosphorylated. A ligandum kötődése konformációs változást indukálhat, ami elősegíti a receptor dimer kialakulását transzfoszforiláció a két receptor lánc között autofoszforiláció.")
12
Receptor tirozin kinázok és a ras
Activation of Ras following binding of a hormone (e.g., EGF) to an RTK. The adapter protein GRB2 binds to a specific phosphotyrosine on the activated RTK and to Sos, which in turn interacts with the inactive Ras · GDP. The guanine nucleotide – exchange factor (GEF) activity of Sos then promotes formation of the active Ras · GTP. Note that Ras is tethered to the membrane by a farnesyl anchor. Grb2: growth factor receptor binding protein
 to an RTK. The adapter protein GRB2 binds to a specific phosphotyrosine on the activated RTK and to Sos, which in turn interacts with the inactive Ras · GDP. The guanine nucleotide – exchange factor (GEF) activity of Sos then promotes formation of the active Ras · GTP. Note that Ras is tethered to the membrane by a farnesyl anchor. Grb2: growth factor receptor binding protein.")
13
A ras által aktivált jeltovábbító kináz kaszkád
Raf MEK Kinase cascade that transmits signals downstream from activated Ras protein. In unstimulated cells, most Ras is in the inactive form with bound GDP (top); binding of a growth factor to its RTK leads to formation of the active Ras · GTP (see Figure 20-23). A signaling complex then is assembled downstream of Ras, leading to activation of MAP kinase by phosphorylation of threonine and tyrosine residues separated by a single amino acid. Phosphorylation at both sites is necessary for activation of MAP kinase. Mitogén aktivált protein kinázok (MAPK)
; binding of a growth factor to its RTK leads to formation of the active Ras · GTP (see Figure 20-23). A signaling complex then is assembled downstream of Ras, leading to activation of MAP kinase by phosphorylation of threonine and tyrosine residues separated by a single amino acid. Phosphorylation at both sites is necessary for activation of MAP kinase. Mitogén aktivált protein kinázok. (MAPK)")
14
Az inaktiv és az aktív, foszforilált MAP kináz szerkezete
Konformáció változás: Dimerizáció ATP és szubsztrát kötés Kötődés más fehérjékhez Structures of MAP kinase in its inactive, unphosphorylated form (a) and active, phosphorylated form (b). Phosphorylation of MAP kinase by MEK at tyrosine 185 (pY185) and threonine 183 (pT183) leads to a marked conformational change in the phosphorylation lip (red). This change promotes dimerization of MAP kinase and binding of its substrates, ATP and certain proteins.
 and active, phosphorylated form (b). Phosphorylation of MAP kinase by MEK at tyrosine 185 (pY185) and threonine 183 (pT183) leads to a marked conformational change in the phosphorylation lip (red). This change promotes dimerization of MAP kinase and binding of its substrates, ATP and certain proteins.")
15
G fehérjékhez kapcsolódó receptorok (GPCR)
Ligandum kötés Trimer G fehérjékkel kölcsönhatásba lépő helyek A schematic drawing of a G-protein-linked receptor. Receptors that bind protein ligands have a large extracellular ligand-binding domain formed by the part of the polypeptide chain shown in light green. Receptors for small ligands such as adrenaline have small extracellular domains, and the ligand-binding site is usually deep within the plane of the membrane, formed by amino acids from several of the transmembrane segments. The parts of the intracellular domains that are mainly responsible for binding to trimeric G proteins are shown in orange,while those that become phosphorylated during receptor desensitization (discussed later) are shown in red. Foszforilálódó helyek (deszenzitizálódás) Ligandum: hormon, neurotranszmitter, lokális mediátorok (C3a, C5a), fény (rodopszin) és szagérzékelő receptorok ligandum fehérjéi etc.
 are shown in red. Foszforilálódó helyek (deszenzitizálódás) Ligandum: hormon, neurotranszmitter, lokális mediátorok (C3a, C5a), fény (rodopszin) és szagérzékelő receptorok ligandum fehérjéi etc.")
16
A rodopszin sematikus képe a kristályszerkezet alapján

17
Bakteriorodopszin-retinal receptor-ligandum komplex
IL-8 A kemokinek hasonló szerkezetű, kis molekulatömegű fehérjék, melyek G fehérjékhez kötött kemokin receptorokhoz kötődnek. Chemokines are a family of proteins of similar structure that bind to chemokine receptors, themselves part of a large family of G protein-coupled receptors. The chemokines are a large family of small proteins represented here by IL-8 (upper molecule). Each chemokine is thought to have a similar structure. The receptors for the chemokines are members of the large family of seven-span receptors, which also includes the photoreceptor protein rhodopsin and many other receptors. They have seven trans-membrane helices, and all members of this receptor family interact with G proteins. The only solved structure of a seven-span membrane protein is of the bacterial protein bacteriorhodopsin; it is depicted in the lower structure, showing the orientation of the seven trans-membrane helices (blue) with the bound ligand (in this case retinal) in red. Essentially all of this structure would be embedded within the cell membrane. Cylinders represent α helices and arrows β strands. Bakteriorodopszin-retinal receptor-ligandum komplex
. Each chemokine is thought to have a similar structure. The receptors for the chemokines are members of the large family of seven-span receptors, which also includes the photoreceptor protein rhodopsin and many other receptors. They have seven trans-membrane helices, and all members of this receptor family interact with G proteins. The only solved structure of a seven-span membrane protein is of the bacterial protein bacteriorhodopsin; it is depicted in the lower structure, showing the orientation of the seven trans-membrane helices (blue) with the bound ligand (in this case retinal) in red. Essentially all of this structure would be embedded within the cell membrane. Cylinders represent α helices and arrows β strands. Bakteriorodopszin-retinal receptor-ligandum komplex.")
18
Ligandum receptor komplexek sematikus modelljei – eltérő szerkezetű ligandumok kölcsönhatása G-fehérjékhez kötött receptorokkal WNT LH, TSH, FSH Fig. 3. Schematic models of ligand-receptor complexes for structurally diverse ligands interacting with GPCRs. The binding sites for all small molecule ligands in family A receptors are buried in between the 7 TMHs. Competitive antagonists and agonists interact with partially overlapping binding sites in biogenicamine receptors, whereas binding sites for allosteric antagonists have been identified in muscarinic receptors, localized extracellular of the primary binding sites. In many peptide receptors, the binding sites for the native peptides involve residues from both extracellular (N termini and ECLs) and 7TM domains (e.g.,NPY, angiotensin II, and GnRH receptors), whereas the binding site for substance P in the NK1 neurokinin receptor involves only extracellular domains. Glycoprotein hormones (LH, TSH, and FSH) and relaxin peptides interact with LRR region in the N termini and ECLs of their receptors. Thrombin and serine proteases activate PAR1–4 by cleavage of the N terminus. Thereafter, the new N termini activate the receptor as a tethered agonist by interacting with residues in ECL2 (except PAR3). PAR3 is probably activated by a tethered agonist from another membrane protein. Family C contains a large VFM, which contains the binding site for competitive antagonists and agonists in a cleft between 2 domains. A competitive antagonist-bonded complex is shown here. Binding of a small molecule agonist to the binding site leads to cleft closure and trapping of the agonist inside the VFM (closed state conformation), whereas a competitive antagonist stabilize an open state conformation of the VFM. A few noncompetitive antagonists are also known for mGlu receptors, which interact with a buried binding site in the transmembrane domain. The N-terminal CRD in Frizzled receptors contains the binding site for their native ligands, the Wnt proteins.
 and 7TM domains (e.g.,NPY, angiotensin II, and GnRH receptors), whereas the binding site for substance P in the NK1 neurokinin receptor involves only extracellular domains. Glycoprotein hormones (LH, TSH, and FSH) and relaxin peptides interact with LRR region in the N termini and ECLs of their receptors. Thrombin and serine proteases activate PAR1–4 by cleavage of the N terminus. Thereafter, the new N termini activate the receptor as a tethered agonist by interacting with residues in ECL2 (except PAR3). PAR3 is probably activated by a tethered agonist from another membrane protein. Family C contains a large VFM, which contains the binding site for competitive antagonists and agonists in a cleft between 2 domains. A competitive antagonist-bonded complex is shown here. Binding of a small molecule agonist to the binding site leads to cleft closure and trapping of the agonist inside the VFM (closed state conformation), whereas a competitive antagonist stabilize an open state conformation of the VFM. A few noncompetitive antagonists are also known for mGlu receptors, which interact with a buried binding site in the transmembrane domain. The N-terminal CRD in Frizzled receptors contains the binding site for their native ligands, the Wnt proteins.")
19
C5a receptor: opszonizált mikroorganizmus fagocitózisát fokozza
The anaphylotoxin C5a can enhance phagocytosis of opsonized microorganisms. Activation of complement, either by the alternative or the MB-lectin pathway, leads to the deposition of C3b on the surface of the microorganism (left panel). C3b can be bound by the complement receptor CR1 on the surface of phagocytes, but this on its own is insufficient to induce phagocytosis (center panel). Phagocytes also express receptors for the anaphylotoxin C5a, and binding of C5a will now activate the cell to phagocytose microorganisms bound through CR1 (right panel). C5a receptor: opszonizált mikroorganizmus fagocitózisát fokozza
. C3b can be bound by the complement receptor CR1 on the surface of phagocytes, but this on its own is insufficient to induce phagocytosis (center panel). Phagocytes also express receptors for the anaphylotoxin C5a, and binding of C5a will now activate the cell to phagocytose microorganisms bound through CR1 (right panel). C5a receptor: opszonizált mikroorganizmus fagocitózisát fokozza.")
20
LIGANDUMOK: IL-8, C3a, C5a, PAF, CCR5, CXCR4 (HIV co-receptor)
")
21
Molekuláris kölcsönhatások
G fehérjékhez kötött receptorok Molekuláris kölcsönhatások Dynamin b-arrestins c-Src RTKs 20 féle a: Effektorok:

22
Hogyan váltják ki a G fehérjéhez kötött sejtfelszíni receptorok kis, intracelluláris mediátorok keletkezését? Másodlagos hírvivők Two major pathways by which G-protein-linked cell-surface receptors generate small intracellular mediators. In both cases the binding of an extracellular ligand alters the conformation of the cytoplasmic domain of the receptor, causing it to bind to a G protein that activates (or inactivates) a plasma membrane enzyme. In the cyclic AMP (cAMP) pathway the enzyme directly produces cyclic AMP. In the Ca2+ pathway the enzyme produces a soluble mediator (inositol trisphosphate, discussed later) that releases Ca2+ from the endoplasmic reticulum. Like other small intracellular mediators, both cyclic AMP and Ca2+ relay the signal by acting as allosteric effectors: they bind to specific proteins in the cell, altering their conformation and thereby their activity. 10-7 M x A célfehérje konformáció és aktivitás változása cAMP közvetített út Ca2+ közvetített út
 a plasma membrane enzyme. In the cyclic AMP (cAMP) pathway the enzyme directly produces cyclic AMP. In the Ca2+ pathway the enzyme produces a soluble mediator (inositol trisphosphate, discussed later) that releases Ca2+ from the endoplasmic reticulum. Like other small intracellular mediators, both cyclic AMP and Ca2+ relay the signal by acting as allosteric effectors: they bind to specific proteins in the cell, altering their conformation and thereby their activity M 4-5x. A célfehérje konformáció és aktivitás változása. cAMP közvetített út. Ca2+ közvetített út.")
23
A ciklikus AMP, szintézise és lebomlása
The synthesis and degradation of cyclic AMP (cAMP). A pyrophosphatase makes the synthesis of cyclic AMP an irreversible reaction by hydrolyzing the released pyrophosphate dclcircp-dclcircp (not shown). Gyors szintézis, gyors lebomlás
. A pyrophosphatase makes the synthesis of cyclic AMP an irreversible reaction by hydrolyzing the released pyrophosphate dclcircp-dclcircp (not shown). Gyors szintézis, gyors lebomlás.")
24
Emlősőkben hat különféle típusa van Szabályozása: Gs fehérje,
Az adenilil cikláz Adenylyl cyclase. In vertebrates the enzyme usually contains about 1100 amino acid residues and is thought to have two clusters of six transmembrane segments separating two similar cytoplasmic catalytic domains. There are at least six types of this form of adenylyl cyclase in mammals (types I-VI). All of them are stimulated by Gs, but type I, which is found mainly in the brain, is also stimulated by complexes of Ca2+ bound to the Ca2+-binding protein calmodulin 1100 aminosav, 2 klaszter, Emlősőkben hat különféle típusa van Szabályozása: Gs fehérje, kivétel: I típus: calmodulin
. All of them are stimulated by Gs, but type I, which is found mainly in the brain, is also stimulated by complexes of Ca2+ bound to the Ca2+-binding protein calmodulin aminosav, 2 klaszter, Emlősőkben hat különféle típusa van. Szabályozása: Gs fehérje, kivétel: I típus: calmodulin.")
25
G fehérjéhez kapcsolódó jelátadás modelje
Az adenil cikláz aktivációja A current model of how Gs couples receptor activation to adenylyl cyclase activation. As long as the extracellular signaling ligand remains bound, the receptor protein can continue to activate molecules of Gs protein, thereby amplifying the response. More important, an αs can remain active and continue to stimulate a cyclase molecule for many seconds after the signaling ligand dissociates from the receptor, providing even greater amplification.

26
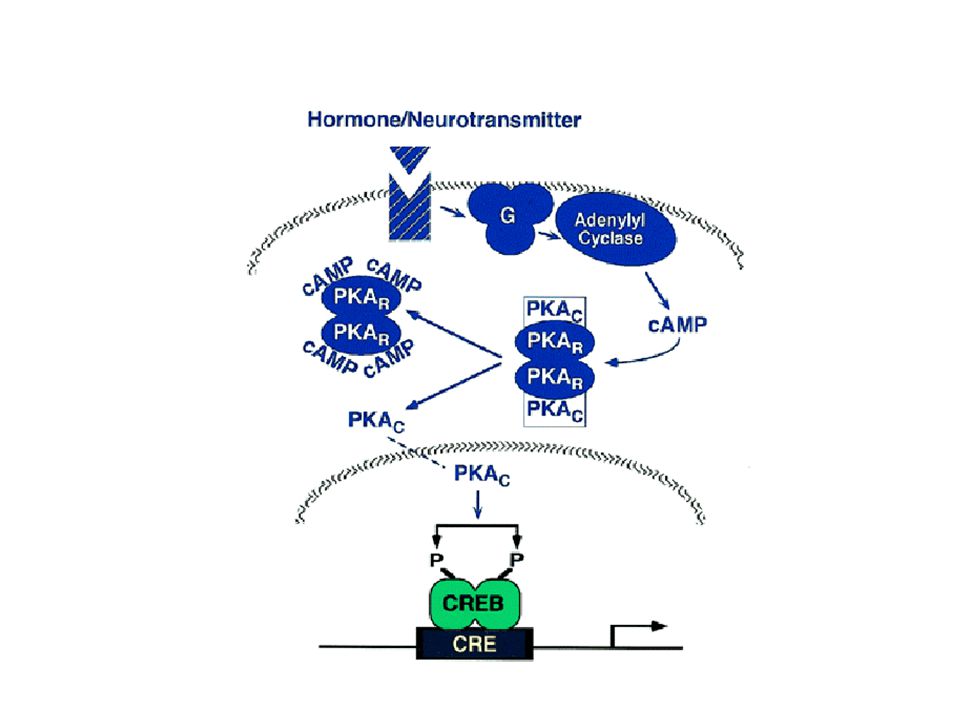
A cAMP-től függő protein kináz (A kináz) aktiválódása
(1968, Walsh and Krebs) The activation of cyclic-AMP-dependent protein kinase (A-kinase). The binding of cyclic AMP to the regulatory subunits induces a conformational change, causing these subunits to dissociate from the complex, thereby activating the catalytic subunits. Each regulatory subunit has two cyclic-AMP-binding sites, and the release of the catalytic subunits requires the binding of more than two cyclic AMP molecules to the tetramer. This greatly sharpens the response of the kinase to changes in cyclic AMP concentration, as we discuss later. There are at least two types of A-kinase in most mammalian cells: type I is mainly in the cytosol, whereas type II is bound via its regulatory subunit to the plasma membrane, nuclear membrane, and microtubules. In both cases, however, once the catalytic subunits are freed and active, they can migrate into the nucleus (where they can phosphorylate gene regulatory proteins), while the regulatory subunits remain in the cytoplasm. The three-dimensional structure of the protein kinase domain of the A-kinase catalytic subunit is shown in Figure 5-12 (next). I típus: citoplazma II típus: membránhoz kötött regulátor alegység Vándorlás a magba -> gén reguláció (pl. CREB foszforiláció) Szubsztrát specificitás: RRxS/T
 The activation of cyclic-AMP-dependent protein kinase (A-kinase). The binding of cyclic AMP to the regulatory subunits induces a conformational change, causing these subunits to dissociate from the complex, thereby activating the catalytic subunits. Each regulatory subunit has two cyclic-AMP-binding sites, and the release of the catalytic subunits requires the binding of more than two cyclic AMP molecules to the tetramer. This greatly sharpens the response of the kinase to changes in cyclic AMP concentration, as we discuss later. There are at least two types of A-kinase in most mammalian cells: type I is mainly in the cytosol, whereas type II is bound via its regulatory subunit to the plasma membrane, nuclear membrane, and microtubules. In both cases, however, once the catalytic subunits are freed and active, they can migrate into the nucleus (where they can phosphorylate gene regulatory proteins), while the regulatory subunits remain in the cytoplasm. The three-dimensional structure of the protein kinase domain of the A-kinase catalytic subunit is shown in Figure 5-12 (next). I típus: citoplazma. II típus: membránhoz kötött regulátor alegység. Vándorlás a magba -> gén reguláció (pl. CREB foszforiláció) Szubsztrát specificitás: RRxS/T.")
27
Activation loop, catalytic loop, substrate binding site

29
A glikogén lebomlás cAMP - PKA által közvetített módja vázizom sejtekben I
Adrenalin PKA közvetített foszfatáz gátlás Foszfatázok: foszforiláció gátlás The stimulation of glycogen breakdown by cyclic AMP in skeletal muscle cells. The binding of cyclic AMP to A-kinase activates this enzyme to phosphorylate and thereby activate phosphorylase kinase, which in turn phosphorylates and activates glycogen phosphorylase, the enzyme that breaks down glycogen. The A-kinase also directly and indirectly increases the phosphorylation of glycogen synthase, which inhibits the enzyme, thereby shutting off glycogen synthesis (not shown). Ser/Thr foszfatáz I, IIA, IIB Foszforiláz kináz
. Ser/Thr foszfatáz I, IIA, IIB. Foszforiláz kináz.")
30
A glikogén lebontás PKA általi szabályozása II:
fehérje foszfatáz I gátlásán keresztül The role of protein phosphatase-I in the regulation of glycogen metabolism by cyclic AMP. Cyclic AMP inhibits protein phosphatase-I, which would otherwise oppose the phosphorylation reactions stimulated by cyclic AMP. It does so by activating A-kinase to phosphorylate a phosphatase inhibitor protein, which then binds to and inhibits protein phosphatase-I. Sokszoros hatás !

31
7x membránon áthaladó receptorok jeltovábbítása-
a legnagyobb receptor család! kis afinitású állapot nagy afinitású állapot A humán genom kb. 1%-a G fehérjékhez kötött receptorokat kódol Figure 1 | Classical examples of seven-transmembrane (7TM)-receptor signalling. In the absence of agonist, 7TM receptors such as the 2 adrenergic receptor ( 2-AR) are in the low-affinity state. After agonist binding, a transient high-affinity complex of agonist, activated receptor and G protein is formed. GDP is released from the G protein and is replaced by GTP. This leads to dissociation of the G-protein complexes into subunits and dimers, which both activate several effectors. G s, for instance, activates adenylyl cyclase, which leads to an increase in cyclic AMP (cAMP). This increase in cAMP in turn activates protein kinase A (PKA), which is a serine/threonine kinase that phosphorylates many different substrates, including 7TM receptors, other kinases and transcription factors. 3; (2002)
-receptor signalling. In the absence of agonist, 7TM receptors such as the 2 adrenergic receptor ( 2-AR) are in the low-affinity state. After agonist binding, a transient high-affinity complex of agonist, activated receptor and G protein is formed. GDP is released from the G protein and is replaced by GTP. This leads to dissociation of the G-protein complexes into subunits and dimers, which both activate several effectors. G s, for instance, activates adenylyl cyclase, which leads to an increase in cyclic AMP (cAMP). This increase in cAMP in turn activates protein kinase A (PKA), which is a serine/threonine kinase that phosphorylates many different substrates, including 7TM receptors, other kinases and transcription factors. 3; (2002)")
32
A 7-transzmembrán receptorok Erk/MAPK aktiváló utjai
Apoptosis signal-regulating kinase (ASK) 7TM receptors activate the ERK/MAPK cascade by several different pathways Mitogen-activated protein kinase (MAPK) cascades are phosphorelay systems in which a MAPK kinase kinase (MAPKKK) is phosphorylated and, in turn, phosphorylates a MAPK kinase (MAPKK), which leads to the phosphorylation of a MAPK. This activated MAPK, a serine/threonine kinase, is then shuttled into the nucleus, where it phosphorylates and regulates its target proteins, ultimately leading to gene transcription and cell proliferation. There are three families of mammalian MAPKs: the extracellular signal-regulated kinases 1/2 (ERK1 and ERK2); the c-Jun amino-terminal kinases (JNKs); and the p38 MAPKs. Seven-transmembrane (7TM) receptors are known to activate each of the different families of MAPKs through many different pathways, including both G - and -dependent mechanisms, and a particular receptor can activate MAPKs through different pathways, depending on the cell context. 7TM-receptor-activation of MAPKs is conserved from yeast to mammals. In yeast, activation of MAPKs is involved in mating. Activation of the ERK cascades is important for proliferation, development and differentiation. Activation of the JNK and p38 MAPK cascades are important for development, inflammation and apoptosis.
 7TM receptors activate the ERK/MAPK cascade by several different pathways Mitogen-activated protein kinase (MAPK) cascades are phosphorelay systems in which a MAPK kinase kinase (MAPKKK) is phosphorylated and, in turn, phosphorylates a MAPK kinase (MAPKK), which leads to the phosphorylation of a MAPK. This activated MAPK, a serine/threonine kinase, is then shuttled into the nucleus, where it phosphorylates and regulates its target proteins, ultimately leading to gene transcription and cell proliferation. There are three families of mammalian MAPKs: the extracellular signal-regulated kinases 1/2 (ERK1 and ERK2); the c-Jun amino-terminal kinases (JNKs); and the p38 MAPKs. Seven-transmembrane (7TM) receptors are known to activate each of the different families of MAPKs through many different pathways, including both G - and -dependent mechanisms, and a particular receptor can activate MAPKs through different pathways, depending on the cell context. 7TM-receptor-activation of MAPKs is conserved from yeast to mammals. In yeast, activation of MAPKs is involved in mating. Activation of the ERK cascades is important for proliferation, development and differentiation. Activation of the JNK and p38 MAPK cascades are important for development, inflammation and apoptosis.")
33
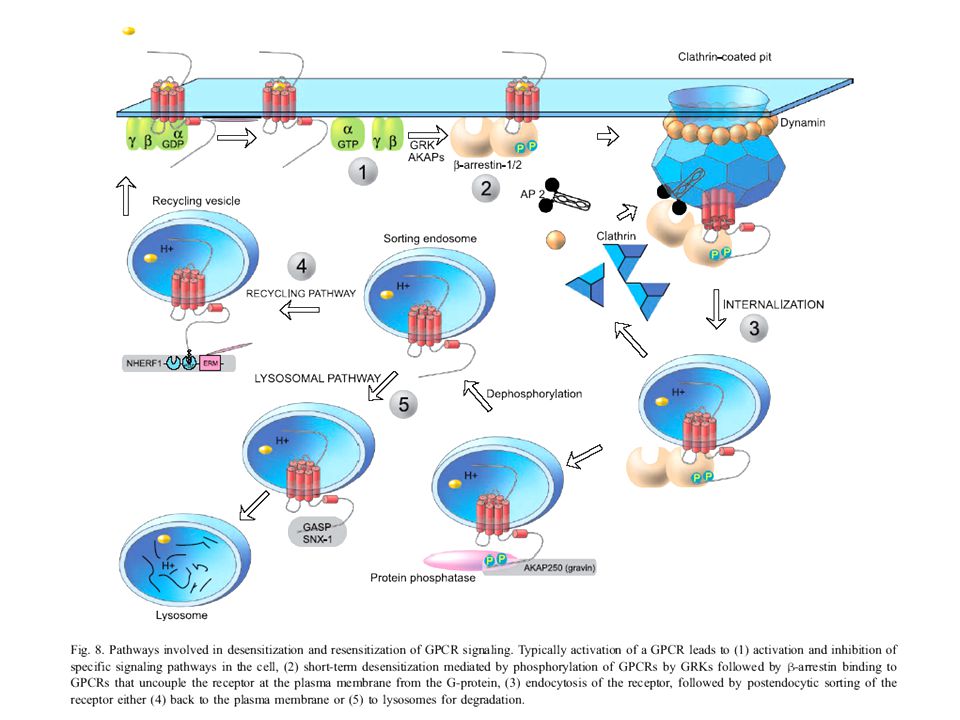
Szabályozás: G fehérjékhez kapcsolt receptorok (GPCR) foszforilációtól független deszenzitizálódása Fig. 1. Potential mechanisms of phosphorylation-independent desensitization of GPCRs. Agonist binding promotes the interaction of three classes of proteins to GPCRs: G proteins, GRKs, and arrestins. GRK interaction mediates uncoupling of receptor and G protein (desensitization) through GPCR phosphorylation, physical association with the receptor, or direct association with and inhibition of Gαq/11. Arrestin interaction with GPCRs also directly inhibits receptor-G protein uncoupling, whereas receptor trafficking mediates various processes, including desensitization, resensitization, and down-regulation. A, agonist. GPCR kináz
 through GPCR phosphorylation, physical association with the receptor, or direct association with and inhibition of Gαq/11. Arrestin interaction with GPCRs also directly inhibits receptor-G protein uncoupling, whereas receptor trafficking mediates various processes, including desensitization, resensitization, and down-regulation. A, agonist. GPCR kináz.")
35
A beta-arrestin egy adaptor/scaffolding (állvány) fehérje a MAPK kaszkád fehérjéi számára – az aktív MAPK-t a citoplazmában tartja Figure 3 | Beta-arrestin scaffolding of mitogen-activated protein kinase (MAPK) cascades. -arrestins function as adaptor/scaffolding molecules, which facilitate the activation of two MAPK cascades — the extracellular-signal regulated kinase (ERK)/MAPK cascade and the c-Jun-amino terminal kinase-3 (JNK3) cascade. a | After G-protein-regulated kinase (GRK)-dependent phosphorylation of the 7TM receptor, -arrestin is recruited to the receptor (b), and brings with it the components of the MAPK cascade; the MAPK kinase kinase (Raf), the MAPK kinase (MEK1), and the MAPK (ERK). c | This association facilitates the activation of ERK/MAPK and targets the active MAPK to the cytosol, thereby decreasing nuclear ERK/MAPK signalling. -arrestin-dependent scaffolding of the JNK3 is highly analogous — -arrestin both facilitates JNK3 activation and leads to the agonist-dependent co-localization of the receptor, -arrestin and the components of the JNK3 cascade in endosomal vesicles.
 cascades. -arrestins function as adaptor/scaffolding molecules, which facilitate the activation of two MAPK cascades — the extracellular-signal regulated kinase (ERK)/MAPK cascade and the c-Jun-amino terminal kinase-3 (JNK3) cascade. a | After G-protein-regulated kinase (GRK)-dependent phosphorylation of the 7TM receptor, -arrestin is recruited to the receptor (b), and brings with it the components of the MAPK cascade; the MAPK kinase kinase (Raf), the MAPK kinase (MEK1), and the MAPK (ERK). c | This association facilitates the activation of ERK/MAPK and targets the active MAPK to the cytosol, thereby decreasing nuclear ERK/MAPK signalling. -arrestin-dependent scaffolding of the JNK3 is highly analogous — -arrestin both facilitates JNK3 activation and leads to the agonist-dependent co-localization of the receptor, -arrestin and the components of the JNK3 cascade in endosomal vesicles.")
36
A G fehérjéhez kötött receptorok másik jeltovábbító rendszere:
A foszfolipáz C b (PLC b) által közvetített jelátvitel
 által közvetített jelátvitel.")
37
Inozitol foszfolipidek (foszfoinozitidek)
Inositol phospholipids (phosphoinositides). The polyphosphoinositides (PIP and PIP2) are produced by the phosphorylation of phosphatidylinositol (PI). Although all three inositol phospholipids may be broken down in the signaling response, it is the breakdown of PIP2 that is most critical, even though it is the least abundant, constituting less than 10% of the total inositol lipids and less than 1% of the total phospholipids. PLC
. The polyphosphoinositides (PIP and PIP2) are produced by the phosphorylation of phosphatidylinositol (PI). Although all three inositol phospholipids may be broken down in the signaling response, it is the breakdown of PIP2 that is most critical, even though it is the least abundant, constituting less than 10% of the total inositol lipids and less than 1% of the total phospholipids. PLC.")
38
A PIP2 hidrolízise The hydrolysis of PIP2. Two intracellular mediators are produced when PIP2 is hydrolyzed: inositol trisphosphate (IP3), which diffuses through the cytosol and releases Ca2+ from the ER, and diacylglycerol, which remains in the membrane and helps activate the enzyme protein kinase C (see below). There are at least three classes of phospholipase C - β, γ, and delta - and it is the β class that is activated by G-protein-linked receptors. We shall see later that the γ class is activated by a second class of receptors, called receptor tyrosine kinases,that activate the inositol-phospholipid signaling pathway without an intermediary G protein.
, which diffuses through the cytosol and releases Ca2+ from the ER, and diacylglycerol, which remains in the membrane and helps activate the enzyme protein kinase C (see below). There are at least three classes of phospholipase C - β, γ, and delta - and it is the β class that is activated by G-protein-linked receptors. We shall see later that the γ class is activated by a second class of receptors, called receptor tyrosine kinases,that activate the inositol-phospholipid signaling pathway without an intermediary G protein.")
39
Az inozitolfoszfolipid által közvetített két jelpálya
The two branches of the inositol phospholipid pathway. The activated receptor binds to a specific trimeric G protein (Gq), causing the α subunit to dissociate and activate phospholipase C-β, which cleaves PIP2 to generate IP3 and diacylglycerol. The diacylglycerol (together with bound Ca2+ and phosphatidylserine - not shown) activates C-kinase. Both phospholipase C-β and C-kinase are water-soluble enzymes that translocate from the cytosol to the inner face of the plasma membrane in the process of being activated. The effects of IP3 can be mimicked experimentally in intact cells by treatment with Ca2+ ionophores, while the effects of diacylglycerol can be mimicked by treatment with phorbol esters, which bind to C-kinase and activate it. Áthelyeződés a citoszólból a membránhoz
, causing the α subunit to dissociate and activate phospholipase C-β, which cleaves PIP2 to generate IP3 and diacylglycerol. The diacylglycerol (together with bound Ca2+ and phosphatidylserine - not shown) activates C-kinase. Both phospholipase C-β and C-kinase are water-soluble enzymes that translocate from the cytosol to the inner face of the plasma membrane in the process of being activated. The effects of IP3 can be mimicked experimentally in intact cells by treatment with Ca2+ ionophores, while the effects of diacylglycerol can be mimicked by treatment with phorbol esters, which bind to C-kinase and activate it. Áthelyeződés a citoszólból a membránhoz.")
40
A PKC által indukált génátírás kétféle módja
I. PKC-közvetített jelpálya A PKC által indukált génátírás kétféle módja Two intracellular pathways by which activated C-kinase can activate the transcription of specific genes. In one ( red arrows) C-kinase activates a phosphorylation cascade that leads to the phosphorylation of a pivotal protein kinase called MAP-kinase (discussed later), which in turn phosphorylates and activates the gene regulatory protein Elk-1. Elk-1 is bound to a short DNA sequence (called serum response element, SRE) in association with another DNA-binding protein (called serum response factor, SRF) . In the other pathway ( green arrows) C-kinase activation leads to the phosphorylation of Ik-B, which releases the gene regulatory protein NF-kB so that it can migrate into the nucleus and activate the transcription of specific genes.
 C-kinase activates a phosphorylation cascade that leads to the phosphorylation of a pivotal protein kinase called MAP-kinase (discussed later), which in turn phosphorylates and activates the gene regulatory protein Elk-1. Elk-1 is bound to a short DNA sequence (called serum response element, SRE) in association with another DNA-binding protein (called serum response factor, SRF) . In the other pathway ( green arrows) C-kinase activation leads to the phosphorylation of Ik-B, which releases the gene regulatory protein NF-kB so that it can migrate into the nucleus and activate the transcription of specific genes.")
41
Calcium ionok által közvetített jelek
II. Calcium ionok által közvetített jelek

42
Szabad Ca2+ koncentráció szabályozása a citoplazmában:
Calcium pumpák, Ca2+ kötő fehérjék 10-3 M Controls on cytosolic Ca2+. The schematic drawing shows the main ways in which cells maintain a very low concentration of free Ca2+ in the cytosol in the face of high concentrations of Ca2+ in the extracellular fluid. Ca2+ is actively pumped out of the cytosol to the cell exterior (A) and into the ER (B). In addition, various molecules in the cell bind free Ca2+ tightly. Mitochondria can also pump Ca2+ out of the cytosol, but they do so efficiently only when Ca2+ levels are extremely high - usually as a result of cell damage. 10-7 M Sejt károsodása, magas Ca2+ koncentráció esetén
 and into the ER (B). In addition, various molecules in the cell bind free Ca2+ tightly. Mitochondria can also pump Ca2+ out of the cytosol, but they do so efficiently only when Ca2+ levels are extremely high - usually as a result of cell damage M. Sejt károsodása, magas Ca2+ koncentráció esetén.")
43
A Ca2+ sejtbe jutásának két módja
Two common pathways by which Ca2+ can enter the cytosol in response to extracellular signals. In (A) Ca2+ enters a nerve terminal from the extracellular fluid through voltage-gated Ca2+ channels when the nerve terminal membrane is depolarized by an action potential. In (B) the binding of an extracellular signaling molecule to a cell-surface receptor generates inositol trisphosphate, which stimulates the release of Ca2+from the ER.
 Ca2+ enters a nerve terminal from the extracellular fluid through voltage-gated Ca2+ channels when the nerve terminal membrane is depolarized by an action potential. In (B) the binding of an extracellular signaling molecule to a cell-surface receptor generates inositol trisphosphate, which stimulates the release of Ca2+from the ER.")
44
Ca 2+-től függő fehérjék A Ca2+ -calmodulin szerkezete
Asp, Glu oldalláncok kötik The structure of Ca2+/calmodulin based on x-ray diffraction and NMR studies. (A) The molecule has a "dumbbell" shape, with two globular ends connected by a long, exposed α helix. Each end has two Ca2+-binding domains, each with a loop of 12 amino acid residues in which aspartic acid and glutamic acid side chains form ionic bonds with Ca2+. The two Ca2+-binding sites in the carboxyl-terminal part of the molecule have a tenfold higher affinity for Ca2+ than those in the amino-terminal part. In solution the molecule is flexible, displaying a range of forms, from extended (as shown) to more compact. (B) The structural changes in Ca2+/calmodulin that occurs when it binds to a target protein (in this example a peptide that consists of the Ca2+/calmodulin-binding domain of a Ca2+/calmodulin-dependent protein kinase [myosin light-chain kinase, discussed below]). Note that the Ca2+/calmodulin has "jack-knifed" to surround the peptide. (A, based on x-ray crystallographic data from Y.S. Babu et al., Nature 315:37-40, © 1985 Macmillan Magazines Ltd.; B, based on x-ray crystallographic data from W.E. Meador, A.R. Means, and F.A. Quiocho, Science 257: , 1992, and on NMR data from M. Ikura et al., Science 256: , 1992.
 The molecule has a dumbbell shape, with two globular ends connected by a long, exposed α helix. Each end has two Ca2+-binding domains, each with a loop of 12 amino acid residues in which aspartic acid and glutamic acid side chains form ionic bonds with Ca2+. The two Ca2+-binding sites in the carboxyl-terminal part of the molecule have a tenfold higher affinity for Ca2+ than those in the amino-terminal part. In solution the molecule is flexible, displaying a range of forms, from extended (as shown) to more compact. (B) The structural changes in Ca2+/calmodulin that occurs when it binds to a target protein (in this example a peptide that consists of the Ca2+/calmodulin-binding domain of a Ca2+/calmodulin-dependent protein kinase [myosin light-chain kinase, discussed below]). Note that the Ca2+/calmodulin has jack-knifed to surround the peptide. (A, based on x-ray crystallographic data from Y.S. Babu et al., Nature 315:37-40, © 1985 Macmillan Magazines Ltd.; B, based on x-ray crystallographic data from W.E. Meador, A.R. Means, and F.A. Quiocho, Science 257: , 1992, and on NMR data from M. Ikura et al., Science 256: ,")
45
A CaM kináz aktiválódása
a alegysége (agyban) The activation of CaM-kinase II. The enzyme is a protein complex of about 12 subunits. The subunits are of four homologous kinds (α, β, γ, and delta), which are expressed in different proportions in different cell types. Only the α subunit, which is expressed only in the brain, is shown (in gray). In the absence of Ca2+/calmodulin the enzyme is inactive as the result of an interaction between the inhibitory domain and the catalytic domain. The binding of Ca2+/calmodulin alters the conformation of the protein, allowing the catalytic domain to phosphorylate the inhibitory domain of neighboring subunits in the complex, as well as other proteins in the cell (not shown). The autophosphorylation of the enzyme complex (by mutual phosphorylation of its subunits) prolongs the activity of the enzyme in two ways: (1) it traps the bound Ca2+/calmodulin so that it does not dissociate from the enzyme complex until cytosolic Ca2+ levels return to basal values for at least 10 seconds (not shown); (2) it converts the enzyme to a Ca2+-independent form so that the kinase remains active even after the Ca2+/calmodulin dissociates from it. Activity continues until the autophosphorylation process is overwhelmed by a protein phosphatase. Ca2+ koncentráció csökken
 The activation of CaM-kinase II. The enzyme is a protein complex of about 12 subunits. The subunits are of four homologous kinds (α, β, γ, and delta), which are expressed in different proportions in different cell types. Only the α subunit, which is expressed only in the brain, is shown (in gray). In the absence of Ca2+/calmodulin the enzyme is inactive as the result of an interaction between the inhibitory domain and the catalytic domain. The binding of Ca2+/calmodulin alters the conformation of the protein, allowing the catalytic domain to phosphorylate the inhibitory domain of neighboring subunits in the complex, as well as other proteins in the cell (not shown). The autophosphorylation of the enzyme complex (by mutual phosphorylation of its subunits) prolongs the activity of the enzyme in two ways: (1) it traps the bound Ca2+/calmodulin so that it does not dissociate from the enzyme complex until cytosolic Ca2+ levels return to basal values for at least 10 seconds (not shown); (2) it converts the enzyme to a Ca2+-independent form so that the kinase remains active even after the Ca2+/calmodulin dissociates from it. Activity continues until the autophosphorylation process is overwhelmed by a protein phosphatase. Ca2+ koncentráció csökken.")
46
A receptor által kiváltott jel erősítése
Amplification in a ligand-induced catalytic cascade. The first amplification step requires that the signaling ligand remain bound to the receptor long enough for the complex to activate many Gs molecules; in many cases the ligand will dissociate too quickly for this amplification to occur.

47
Kis G fehérjék: ras, rac, rho

48
GTP-kötött ras féléletidő: 1-5 óra, saját GTPáz aktivitás.
Ras-GAP: negatív regulátor, fokozza „ „ ” ” Mutáció Konstitutív aktivitás Tumor Ras: sejtosztódás, differenciáció Rho: actin-citoszeleton kapcsolat, sejtosztódást befoly.

49
A SOS molekula és kapcsolatai
The Two Hats of SOS Anjaruwee Nimnual and Dafna Bar-Sagi* (Science’s STKE 2002, 145 pe36) Rac cserefaktor aktivitás: Fig. 1. Molecular organization of the SOS-E3b1-Eps8 tricomplex. E3b1 mediates the interaction of SOS with Eps8 through an SH3-dependent binding to the first of four proline-rich (P-rich) motifs in the COOH terminal region of SOS, and proline-rich-dependent binding to the SH3 domain of Eps8. The effector region of Eps8 confers Rac exchange activity on SOS and targets the protein complex to actin filaments. Dotted lines denote confirmed interactions; the dashed line denotes a presumptive interaction. Dbl homológ domén pleckstrin homológ domén Rho GTPase cserefaktorokra jellemző „Signature motif” Katalitikusan aktív domén Ras cserefaktorokra jellemző
 Rac cserefaktor aktivitás: Fig. 1. Molecular organization of the SOS-E3b1-Eps8 tricomplex. E3b1 mediates the interaction of SOS. with Eps8 through an SH3-dependent binding to the first of four proline-rich (P-rich) motifs in the COOH terminal region of SOS, and proline-rich-dependent binding to the SH3 domain of Eps8. The effector region of Eps8 confers Rac exchange activity on SOS and targets the protein complex to actin filaments. Dotted lines denote confirmed interactions; the dashed line denotes a presumptive interaction. Dbl homológ domén. pleckstrin homológ domén. Rho GTPase cserefaktorokra jellemző „Signature motif Katalitikusan aktív domén. Ras cserefaktorokra jellemző.")
50
A SOS guanin nukleotid cserefaktor által ellátott két funkció:
Ras aktiválás Nyugvó sejt Aktivált sejt Fig. 2. SOS exists in two functional pools. In resting cells, SOS-Grb2 and SOS-E3b1-Eps8 complexes reside in the cytoplasm, and Ras and Rac are in an inactive guanosine diphosphate (GDP)-bound state. Upon growth factor stimulation, the SOS-Grb2 complex translocates to the activated RTK, thereby gaining proximity to its target Ras. The SOS-E3b1-Eps8 complex is recruited to actin filaments, which, in turn, would promote the activation of Rac in the vicinity of its physiological target, the actin cytoskeleton. Rac aktiválás
-bound state. Upon growth factor stimulation, the SOS-Grb2 complex translocates to the activated RTK, thereby gaining proximity to its target Ras. The SOS-E3b1-Eps8 complex is recruited to actin filaments, which, in turn, would promote the activation of Rac in the vicinity of its physiological target, the actin cytoskeleton. Rac aktiválás.")
51
Ión transzport Component Intracellular Concentration (mM)
Extracellular Concentration (mM) Cations Na+ 5-15 145 K+ 140 5 Mg2+ 0.5 1-2 Ca2+ 10-4 H+ 7 x10-5 ( M or pH 7.2) 4x10-5 ( M or pH 7.4) Anions* Cl- 110 *. The cell must contain equal quantities of + and - charges (that is, be electrically neutral). Thus, in addition to Cl-, the cell contains many other anions not listed in this table; in fact, most cellular constituents are negatively charged (HCO3-, PO43-, proteins, nucleic acids, metabolites carrying phosphate and carboxyl groups, etc.). The concentrations of Ca2+ and Mg2+ given are for the free ions. There is a total of about 20 mM Mg2+ and 1-2 mM Ca2+ in cells, but this is mostly bound to proteins and other substances and, in the case of Ca2+, stored within various organelles.
 Cations. Na K Mg Ca H+ 7 x10-5 ( M or pH 7.2) 4x10-5 ( M or pH 7.4) Anions* Cl *. The cell must contain equal quantities of + and - charges (that is, be electrically neutral). Thus, in addition to Cl-, the cell contains many other anions not listed in this table; in fact, most cellular constituents are negatively charged (HCO3-, PO43-, proteins, nucleic acids, metabolites carrying phosphate and carboxyl groups, etc.). The concentrations of Ca2+ and Mg2+ given are for the free ions. There is a total of about 20 mM Mg2+ and 1-2 mM Ca2+ in cells, but this is mostly bound to proteins and other substances and, in the case of Ca2+, stored within various organelles.")
52
Hordozó fehérje által követített transzport
Three types of carrier-mediated transport. The schematic diagram shows carrier proteins functioning as uniports, symports, and antiports. The function of ion channels is to allow specific inorganic ions, mainly Na+, K+, Ca2+, or Cl-, to diffuse rapidly down their electrochemical gradients across the lipid bilayer

53
Na+-K+ ATP-ase The Na+-K+ ATPase. This carrier protein actively pumps Na+ out of and K+ into a cell against their electrochemical gradients. For every molecule of ATP hydrolyzed inside the cell, three Na+ are pumped out and two K+ are pumped in. The specific pump inhibitor ouabain and K+ compete for the same site on the external side of the ATPase. This carrier protein actively pumps Na+ out of and K+ into a cell against their electrochemical gradients. For every molecule of ATP hydrolyzed inside the cell, three Na+ are pumped out and two K+ are pumped in. The specific pump inhibitor ouabain and K+ compete for the same site on the external side of the ATPase.

Hasonló előadás
















>")


