Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Áthelyeződő genetikai elemek
Transzpozon terminológia Inzerciós szekvenciák Transzpozonok kimutatása baktériumokban Bakteriális transzpozonok típusai Transzpozíció (áthelyeződés) Transzpozonok kivágódása A transzpozonok hatása a baktérium genomra Mu fág A transzpozonok és az evolúció
 Transzpozonok kivágódása. A transzpozonok hatása a baktérium genomra. Mu fág. A transzpozonok és az evolúció.")
2
Genetikai, fizikai térképezés eredménye
A gének meghatározott helyen találhatók Bizonyított a genetikai átrendeződés (pl. inverzió, duplikáció stb.) McClintock kimutatja kukoricában az áthelyeződő szabályozó elemeket 30 évvel később a baktériumokban is felfedezik az áthelyeződő elemeket Kis gyakoriságú jelenség ( /generáció) Statikus genomkép átalakul a genom mozgásban van (genome in flux)
 McClintock kimutatja kukoricában az áthelyeződő szabályozó elemeket. 30 évvel később a baktériumokban is felfedezik az áthelyeződő elemeket. Kis gyakoriságú jelenség ( /generáció) Statikus genomkép átalakul a genom mozgásban van (genome in flux)")
3
A genetikai elem mozgása DNS kicserélődéssel jár
Ez nem homológ szakaszok közt jön létre RecA rendszertől független Az áthelyeződés a transzpozíció Az elem neve transzpozon

4
Transzpozon terminológia
A legtöbb baktérium transzpozon könnyen felismerhető géneket tartalmaz Legtöbbször antibiotikum rezisztencia gént hordoz A legtöbb rezisztencia gént hordozó transzpozon elnevezése Tn és egy szám (pl. Tn5) Megegyezés szerint az elnevezés akkor is ez, ha nincs ismert tulajdonság Ha hivatkozunk a hordozott génre azt az általános jelzéssel tesszük pl: Tn1(ampr)
 Megegyezés szerint az elnevezés akkor is ez, ha nincs ismert tulajdonság. Ha hivatkozunk a hordozott génre azt az általános jelzéssel tesszük pl: Tn1(ampr)")
5
Az elsőként felfedezett transzpozonok nem tartalmaztak ismert gént, ezért ezeket inzerciós szekvenciának nevezték el IS elemek IS1, IS2 stb. Van nénány nem standard módon elnevezett transzpozon (pl. Tn1000, amely az F plazmidon található, néven is szerepel) A transzpozonok gyakran valamilyen génen belül találhatók, mutációt okoznak Az elnevezés, lacZ87::Tn3, a 87-es mutáció a lacZ génben, melyet a Tn3 beépülése okozott
 A transzpozonok gyakran valamilyen génen belül találhatók, mutációt okoznak. Az elnevezés, lacZ87::Tn3, a 87-es mutáció a lacZ génben, melyet a Tn3 beépülése okozott.")
6
Inzerciós szekvenciák
Az első áthelyeződő genetikai elemek E. coli-ból származnak (1967) gal- és lac- mutáns kollekciót hoztak létre, melyek különleges tulajdonságokkal rendelkeztek
 gal- és lac- mutáns kollekciót hoztak létre, melyek különleges tulajdonságokkal rendelkeztek.")
7
A mutációk jellegzetességei
1. Nagymértékben poláris mutációk az operon első génjébe térképeződött, de a hátrébb lévő gének terméke nem jelent meg 2. A mutánsoknál nem volt reverzió báziskicserélődéssel, frame-shift mutációval, azaz nem pontmutáció Poláris mutációkról (korábban ismertek) kimutatták, hogy lánc terminációs mutánsok (revertálhatók) 3. Ha egy poláris mutációt hordozó törzsbe plazmidot juttattak át, hasonló poláris mutációk jelentek meg a plazmidon hordozott génekben 4. Fizikai kísérletek kimutatták, hogy a hasonló poláris mutációt hordozó plazmidok nagyobbak voltak, mint az eredeti Azaz inzertált szekvenciákat hordoztak, erről feltételezték, hogy mobilis szakasz
 kimutatták, hogy lánc terminációs mutánsok (revertálhatók) 3. Ha egy poláris mutációt hordozó törzsbe plazmidot juttattak át, hasonló poláris mutációk jelentek meg a plazmidon hordozott génekben. 4. Fizikai kísérletek kimutatták, hogy a hasonló poláris mutációt hordozó plazmidok nagyobbak voltak, mint az eredeti. Azaz inzertált szekvenciákat hordoztak, erről feltételezték, hogy mobilis szakasz.")
8
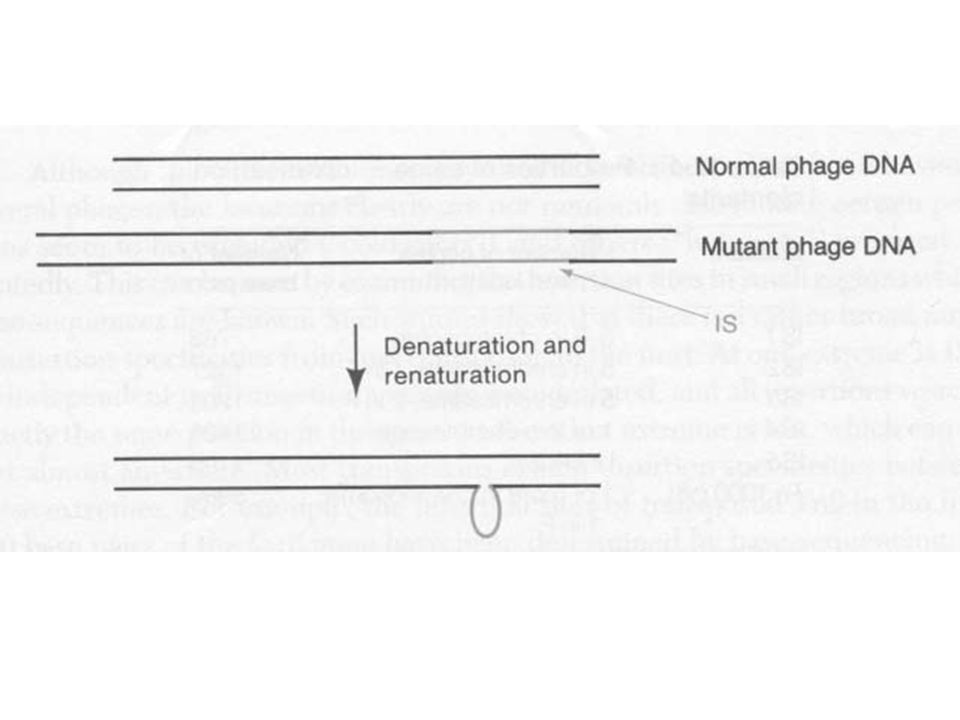
Ha az előző poláris mutációt hordozó törzseket fággal fertőzték, olyan fágokat lehetett izolálni, melyek szintén poláris mutációt hordoztak Fizikai vizsgálatok, heteroduplex analízis kimutatták, hogy a mutáció inzerció Vad típusú és mutáns fág DNS-t denaturálták, majd hibridizálták A vad és mutáns közötti különbség az inzerciós szekvencia, amely a hibridizálás során „kihurkolódott”) Az inzerció nagysága ugyanaz, mint a heteroduplex analízisben a hurok nagysága Ha különböző fág fajokkal fertőzték a poláris mutációt hordozó baktériumtörzset, akkor a mutáns fágok (más faj) hibridizáltak egymással, azaz az inzerciós szekvencia baktériumból származott
 Az inzerció nagysága ugyanaz, mint a heteroduplex analízisben a hurok nagysága. Ha különböző fág fajokkal fertőzték a poláris mutációt hordozó baktériumtörzset, akkor a mutáns fágok (más faj) hibridizáltak egymással, azaz az inzerciós szekvencia baktériumból származott.")
10
A különböző IS elemek különböző nagyságúak
Többféle inzerciós elemet izoláltak, szekvenálás után a következő tulajdonságok derültek ki A poláris mutációt okozó elemek transzkripciós stop szignált és mindhárom frame-ben transzlációs stop szignálokat hordoztak Ezek okozzák a poláris hatást Az IS elemek végein fordítottan ismétlődő (invereted repeat) szekvenciák találhatók pl. AGTC…..GACT, hosszuk bp Sok inzerciós helynél a beépülés iránya változó volt (nem csak egy irányban történt beépülés), target specifikusság A különböző IS elemek különböző nagyságúak Az elemek legalább két ORF-et (nyitott leolvasási keretet) tartalmaznak és a fehérjéket is kimutatták
 szekvenciák találhatók pl. AGTC…..GACT, hosszuk bp. Sok inzerciós helynél a beépülés iránya változó volt (nem csak egy irányban történt beépülés), target specifikusság. A különböző IS elemek különböző nagyságúak. Az elemek legalább két ORF-et (nyitott leolvasási keretet) tartalmaznak és a fehérjéket is kimutatták.")
11
Transzpozonok kimutatása baktériumokban
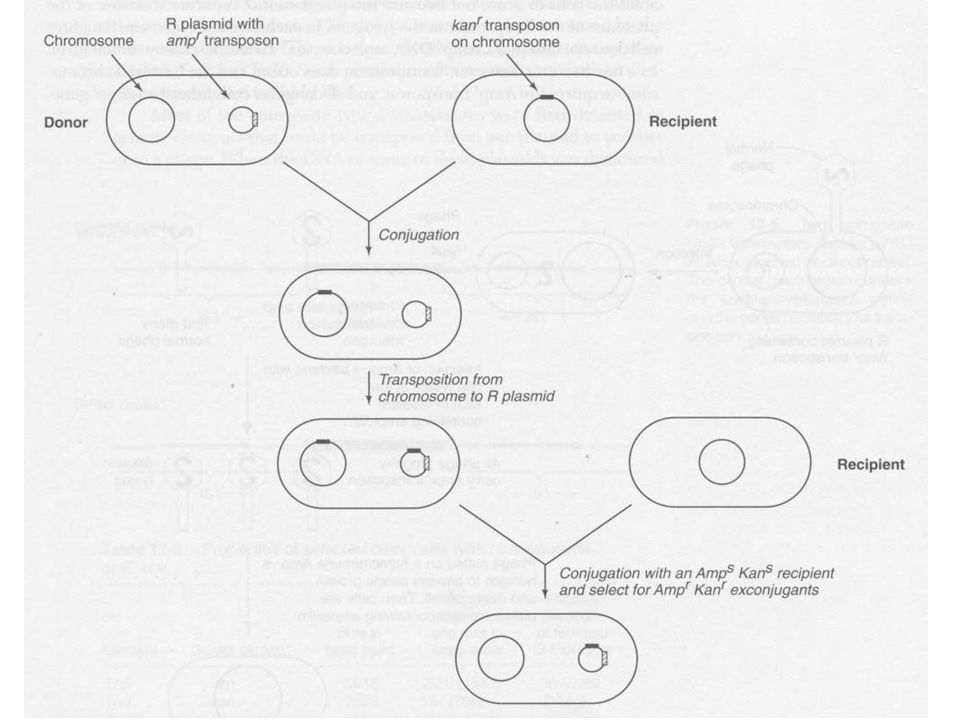
IS elemekkel ellentétben a transzpozonok hordoznak detektálható gént (pl. antibiotikum rezisztencia) Ezekkel követni lehet az áthelyeződést 1. R plazmid Ampr konjugációval RecA- recipiens törzsbe, melyben Kanr-t hordozó transzpozon van Szaporítás során előfordul, hogy a Kanr transzpozon a plazmidra ugrik A plazmidon két transzpozon is van Ampr és Kanr Ezt érzékeny gazdába történő konjugációval mutatták ki
 Ezekkel követni lehet az áthelyeződést. 1. R plazmid Ampr konjugációval RecA- recipiens törzsbe, melyben Kanr-t hordozó transzpozon van. Szaporítás során előfordul, hogy a Kanr transzpozon a plazmidra ugrik. A plazmidon két transzpozon is van Ampr és Kanr. Ezt érzékeny gazdába történő konjugációval mutatták ki.")
13
IS4 pl. 20 galT mutánsból mind egy helyen
A beépülés általában véletlen szerű (random), nincs kitüntetett cél szekvencia Vannak elkerült helyek Preferált helyek (hot spot) IS4 pl. 20 galT mutánsból mind egy helyen Mu fág szinte bárhova inzertálódhat A legtöbb transzpozon a kettő közötti specificitással rendelkezik
, nincs kitüntetett cél szekvencia. Vannak elkerült helyek. Preferált helyek (hot spot) IS4 pl. 20 galT mutánsból mind egy helyen. Mu fág szinte bárhova inzertálódhat. A legtöbb transzpozon a kettő közötti specificitással rendelkezik.")
14
Transzpozíció kimutatása fággal
Lambda fág szaporítás Gazda, recA-, Ampr R plazmid, az R plazmidon transzpozon okozza a rezisztenciát Néhány fág részecske tartalmazza a transzpozont Ezt a keverék lizátumot használták fertőzéshez Amps gazdán

15
Kis MOI és ampicillines szelekcióval a keletkező utód fág mind Ampr (a fágon lévő transzpozon rezisztenciát biztosít a gazdának, a vad típusú fágfertőzés során elpusztul a sejt és fág sem termelődik) A tiszta Ampr fáglizátummal lizogén (homoimmunis) baktériumot fertőztek A lizogén baktériumban nem szaporodik a fág, de a DNS bejut Ampicillines szelekcióval olyan baktériumot izolálhatunk, amelyiknek a kromoszómájába beépült a transzpozon
 baktériumot fertőztek. A lizogén baktériumban nem szaporodik a fág, de a DNS bejut. Ampicillines szelekcióval olyan baktériumot izolálhatunk, amelyiknek a kromoszómájába beépült a transzpozon.")
17
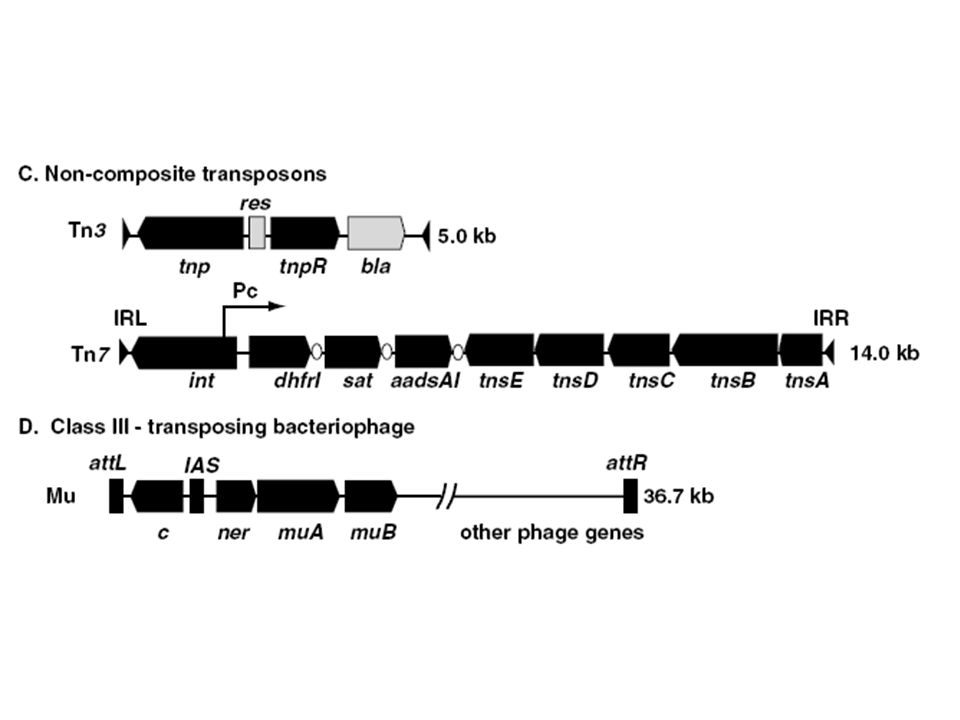
Bakteriális transzpozonok típusai
IS elemek 1. Kompozit transzpozonok 2. Tn3 transzpozon család 3. Áthelyeződő fágok

18
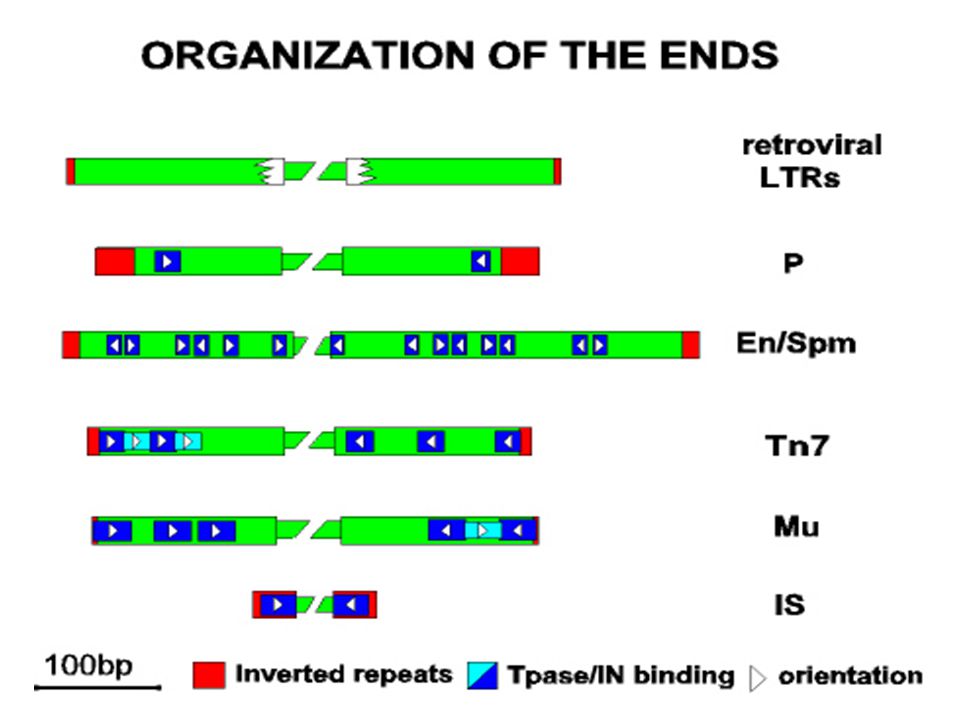
Kompozit (I-es típusú) transzpozonok
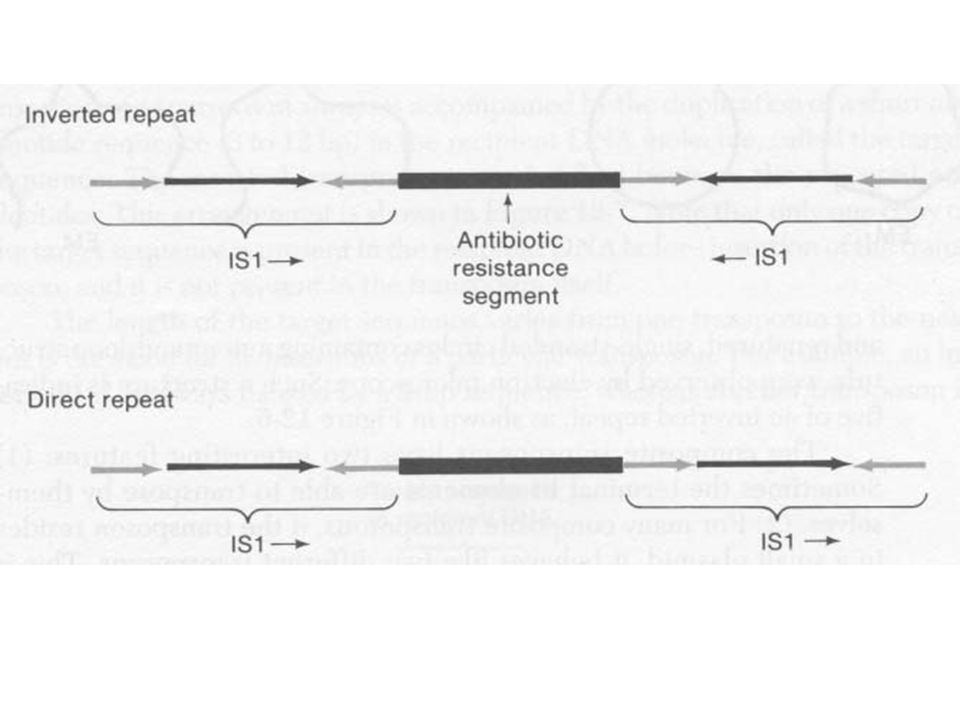
Két azonos / majdnem azonos IS elem határolja Az IS elemek elhelyezkedése fordítottan ismétlődő vagy direkt ismétlődő (inverted, direct repeat) A terminális szekvencia ugyanaz mindkettőnél Az, hogy áthelyeződik vagy sem a transzpozon az IS elemtől (terminális) függ Tn5, Tn9 és Tn10 Elsőként úgy mutatták ki, mint olyan genetikai elemeket, amelyek plazmidról plazmidra ill. plazmidról fágra helyeződnek át Elektronmikroszkóppal denaturálás és gyors renaturálás után stem loop (a fordítottan ismétlődő szakasz miatt)
 A terminális szekvencia ugyanaz mindkettőnél. Az, hogy áthelyeződik vagy sem a transzpozon az IS elemtől (terminális) függ. Tn5, Tn9 és Tn10. Elsőként úgy mutatták ki, mint olyan genetikai elemeket, amelyek plazmidról plazmidra ill. plazmidról fágra helyeződnek át. Elektronmikroszkóppal denaturálás és gyors renaturálás után stem loop (a fordítottan ismétlődő szakasz miatt)")
21
Két érdekes tulajdonság
A terminális IS elemek önmaguk is áthelyeződhetnek Sok kompozit transzpozonnál, ha kis méretű plazmidon van, akkor két különálló transzpozonként is viselkedhet Ha egy plazmidon 2 IS elem van, akkor a plazmid minden része IS elem között van, azaz transzpozon

22
Transzpozíció mechanizmusai

24
Transzpozon családok

28
Tn3 transzpozon család Minden Tn3-hoz hasonló elem két 38 bp nagyságú fordított szekvenciát tartalmaz Nincsen IS-hez hasonló elem rajtuk 3 gént hordoz, -laktamáz (ampicillin rezisztencia), két transzpozícióhoz szükséges gén (transzpozáz, reszolváz)
, két transzpozícióhoz szükséges gén (transzpozáz, reszolváz)")
29
Áthelyeződő fágok Két rokon fág Mu, D108
A transzpozíció az életciklusuk része A fág DNS replikációhoz transzpozíció szükséges

30
Transzpozíció (áthelyeződés)
Célszekvencia duplikálódás a beépülés helyén A transzpozonok és transzpozon célszekvencia kapcsolat szerkezete Replikatív transzpozíció A Tn3 transzpozíciója során a kointegrátum az intermedier A replikatív transzpozíció mechanizmusa Nem replikatív transzpozíció

31
A transzpozíció eredménye, hogy a transzpozon a recipiensbe épül
A transzpozon és inzerciós hely között nincsen homológia recA rendszer sem szükséges hozzá

32
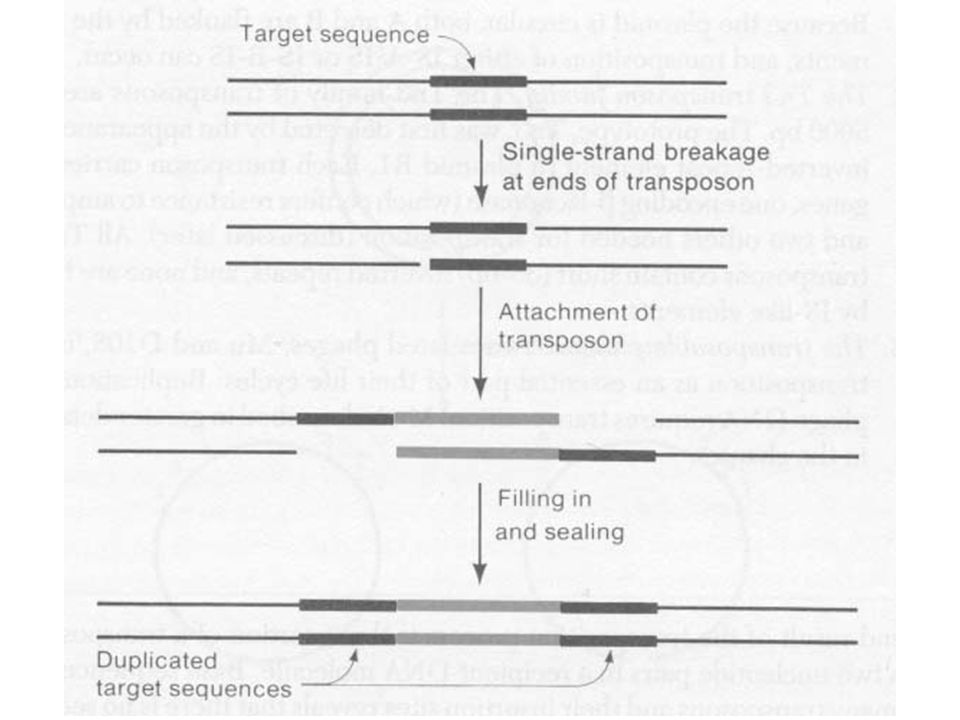
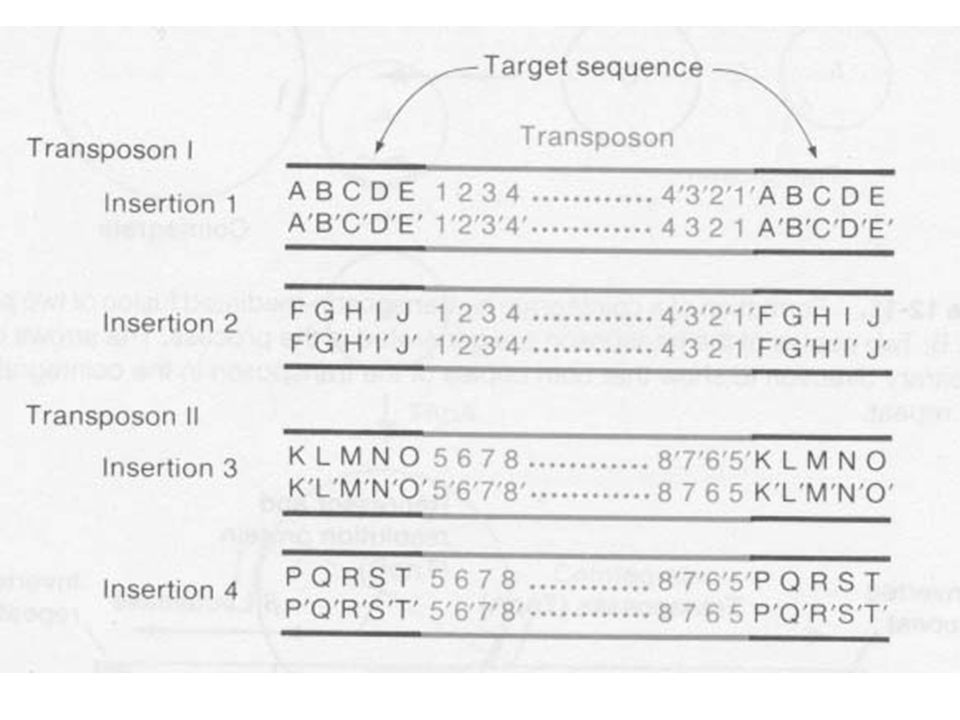
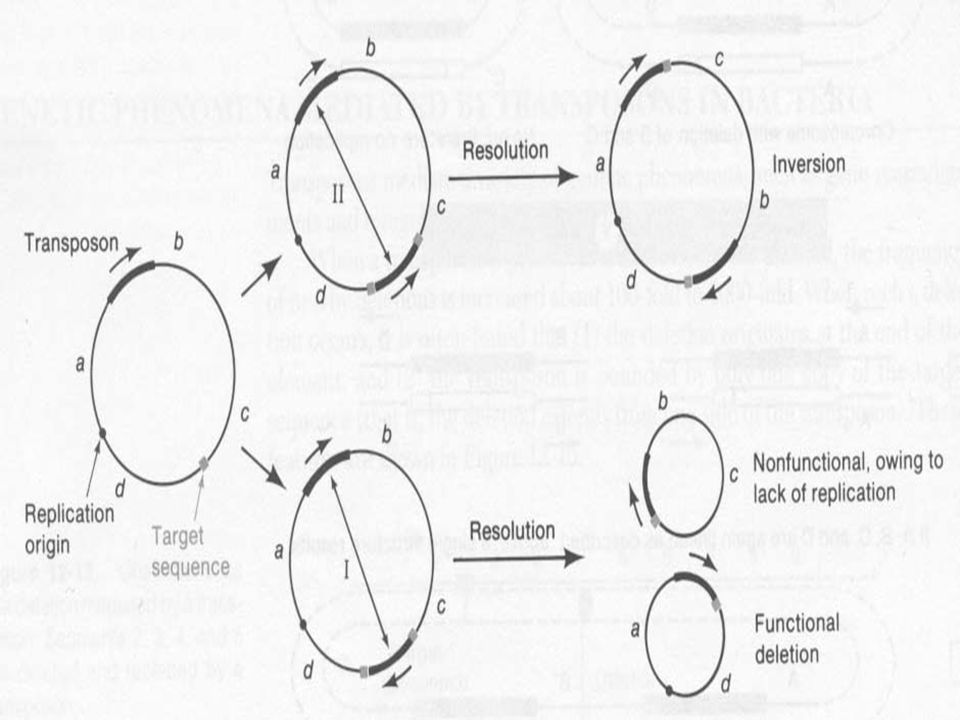
Célszekvencia duplikálódás a beépülés helyén
A beépüléskor a cél szekvencia duplikálódik A transzpozon a két szekvencia között van A célszekvencia hossza transzpozontól függ, de egyfajta transzpozonnál mindig ugyanaz A cél szekvencia transzpozíciónként más és más, de a hossza ugyanaz

34
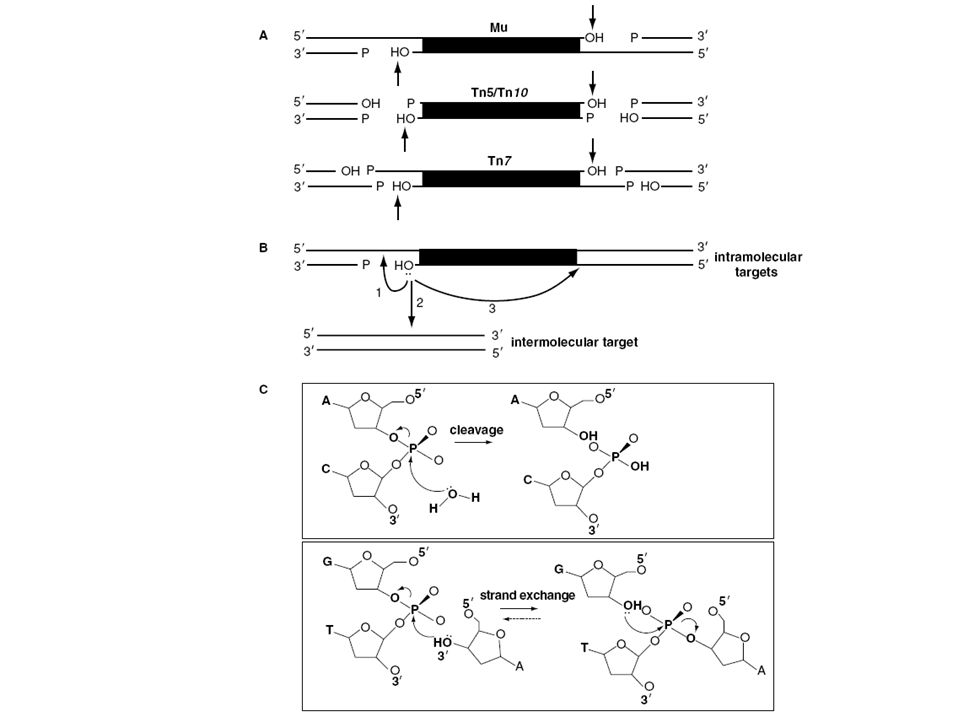
A transzpozáz vágja a DNS-t
„lépcsős” hasítás, vagy két egyszálú nick A transzpozon a szabad véghez kapcsolódik Az egyszálú részek feltöltödnek Ez okozza a cél duplázódását A duplikáció nem feltétele a transzpozíciónak

36
A transzpozonok és transzpozon célszekvencia kapcsolat szerkezete
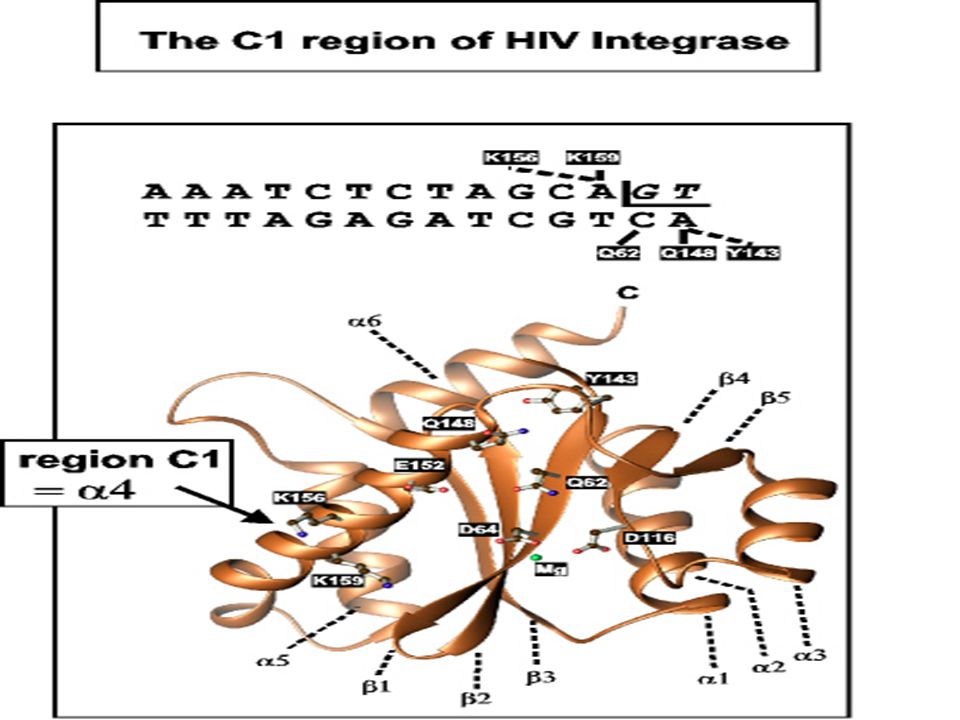
A transzpozonok jól meghatározott végekkel rendelkeznek Minden inzerciónál a transzpozon végei ugyanazok Minden transzpozon saját vég szekvenciával rendelkezik Beépüléskor a transzpozon végek a nick-ekhez kapcsolódnak

37
A transzpozonok végei esszenciálisak a transzpozícióhoz
A legtöbb transzpozonnál a két vég ugyanaz a szekvencia csak fordított orientációban Néhány kivétel A transzpozonok végei esszenciálisak a transzpozícióhoz Báziscsere, inzerció, deléció a terminális szekvenciában megakadályozza a transzpozíciót

39
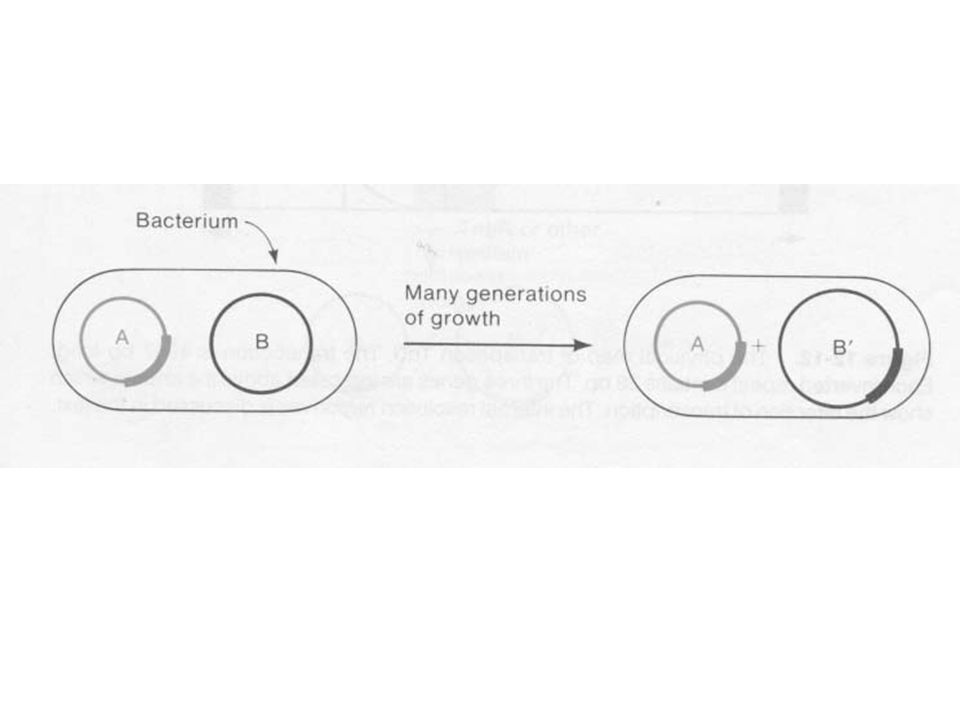
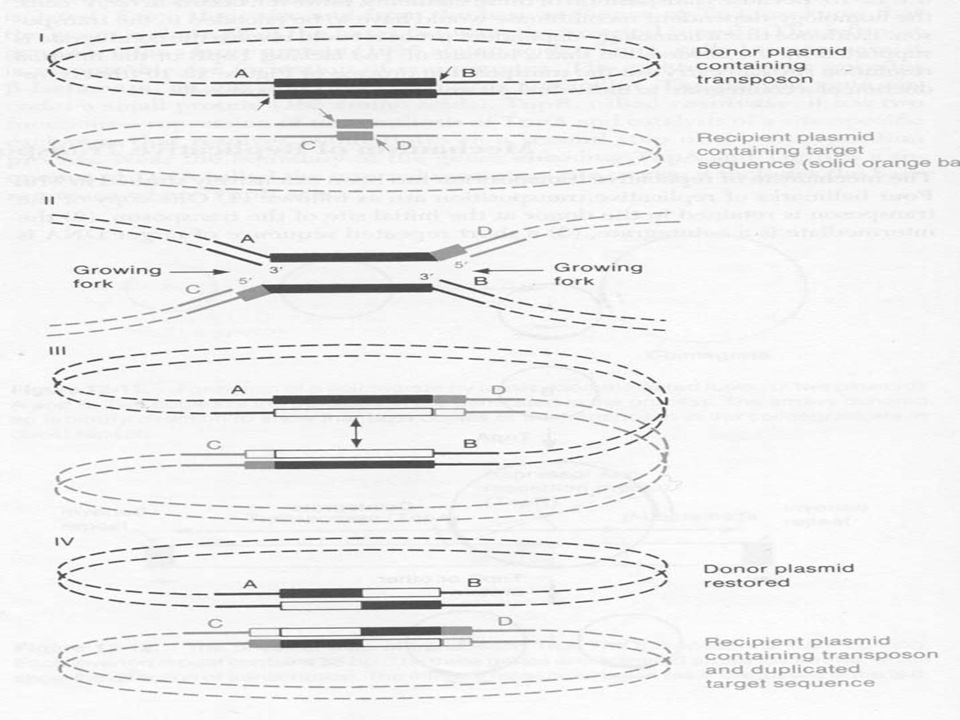
Replikatív transzpozíció
A gazda szaporítása a során (10-7 gyakoriság) a transzpozon a plazmidról átkerülhet egy másik plazmidra Egyes transzpozonoknál ilyenkor mindkét plazmidon megtalálható a transzpozon Az eredeti és egy replikálódott változat Ugyanilyen kis gyakorisággal plazmid kointegrátum keletkezik A két plazmid direkt ismétlődik és két transzpozon van a csatlakozási helynél A nem replikatív transzpozonoknál nincs kointegrátum
 a transzpozon a plazmidról átkerülhet egy másik plazmidra. Egyes transzpozonoknál ilyenkor mindkét plazmidon megtalálható a transzpozon. Az eredeti és egy replikálódott változat. Ugyanilyen kis gyakorisággal plazmid kointegrátum keletkezik. A két plazmid direkt ismétlődik és két transzpozon van a csatlakozási helynél. A nem replikatív transzpozonoknál nincs kointegrátum.")
41
A Tn3 transzpozíciója során a kointegrátum az intermedier
A Tn3 (és a hasonló transzpozonok) replikatív módon helyeződik át A keletkező kointegrátum intermedier a transzpozíció során
 replikatív módon helyeződik át. A keletkező kointegrátum intermedier a transzpozíció során.")
42
A Tn3 szerkezete Két terminális fordított ismétlődő szakasz határolja
3 gén tnpA, transzpozáz, a kointegrátum kialakításáért felelős tnpR, reszolváz, represszálja a TnpA szintézisét, helyspecifikus DNS szál kicserélődést katalizál, azaz a kointegrátumot megoldja res hely, belső megoldó hely -laktamáz, ampicillin bontás

43
Tn3 szerkezete

44
Áthelyeződés két lépésből áll
Mutációk a génekben tnpA, nem képes áthelyeződni, kointegrátumot képezni tnpR, res mutánsok képesek kointegrátumot létrehozni, de nincsen áthelyeződés (tnpR mutáns recA+ törzsben megoldódik) Áthelyeződés két lépésből áll tnpA kointegrátumot hoz létre tnpR katalizálja a res helyen keresztüli kicserélődést Néhány nem Tn3 családba tartozó transzpozonnál nincs reszolváz A kointegrátum homológ rekombinációval is megoldódhat, mert két homológ (transzpozon) szakasz is van a DNS molekulán, nem recA, hanem transzpozon kódolt rekombináz
 Áthelyeződés két lépésből áll. tnpA kointegrátumot hoz létre. tnpR katalizálja a res helyen keresztüli kicserélődést. Néhány nem Tn3 családba tartozó transzpozonnál nincs reszolváz. A kointegrátum homológ rekombinációval is megoldódhat, mert két homológ (transzpozon) szakasz is van a DNS molekulán, nem recA, hanem transzpozon kódolt rekombináz.")
46
A replikatív transzpozíció mechanizmusa
1. A transzpozon egy kópiája megmarad a donor DNS-en az eredeti helyén 2. Az intermedier egy kointegrátum 3. Rövid DNS darab ismétlődik a transzpozon széleinél (target duplikáció) 4. Néhány transzpozonnál szükséges egy belső megoldó hely, vagy homológ szakaszok közti kicserélődés
 4. Néhány transzpozonnál szükséges egy belső megoldó hely, vagy homológ szakaszok közti kicserélődés.")
47
A replikatív transzpozíció indításához két lépés kell
A két egyszálú szakasz összekapcsolódik (anélkül, hogy bázis párosodás lenne) ez a csatlakozási pont Replikációs villa keletkezik a ligálás után
 ez a csatlakozási pont. Replikációs villa keletkezik a ligálás után.")
48
A replikatív transzpozíció lépései
A transzpozáz egy szálat elvág a cél szekvenciában, majd a másik szálat egy kicsit odébb, az eredmény ragadós vég A transzpozon ellentétes végén a komplementer szálon hasítás. Mindkét 3’ vég a recipiens 5’ túlnyúló végéhez kapcsolódik, ez két replikációs villát eredményez A replikáció a cél szekvencia 5’ végénél kezdődik (a ragadós vég szintézisével). Amikor a replikáció befejeződik kointegrátum keletkezik, két transzpozon kópiával Kétszálú kicserélődés történik a transzpozon két kópiája között. Vagy a belső megoldó hely vagy a homológ szakaszok között
. Amikor a replikáció befejeződik kointegrátum keletkezik, két transzpozon kópiával. Kétszálú kicserélődés történik a transzpozon két kópiája között. Vagy a belső megoldó hely vagy a homológ szakaszok között.")
50
Nem replikatív transzpozíció
Konzervatív cut&paste DNS szintézis csak a transzpozon és target közötti rések feltöltéséhez szükséges Kísérletes bizonyíték lacZ mutáns Tn10-en Heteroduplex, az egyik szál mutáns a másik vad típus Replikációkor vagy egyik vagy másik szál Nem replikatív átvitelnél mindkét szál öröklődik

52
Transzpozonok kivágódása
Néha kivágódhatnak Pontos kivágódás (10-6 gyakoriság) Lac mutáns Tn5 Reverzió a transzpozon Km rezisztencia megszűnésével Néhány esetben a Km rezisztencia megmaradt, a transzpozon más helyre transzlokálódott, és a vad lac fenotípus visszaállt
 Lac mutáns Tn5. Reverzió a transzpozon Km rezisztencia megszűnésével. Néhány esetben a Km rezisztencia megmaradt, a transzpozon más helyre transzlokálódott, és a vad lac fenotípus visszaállt.")
53
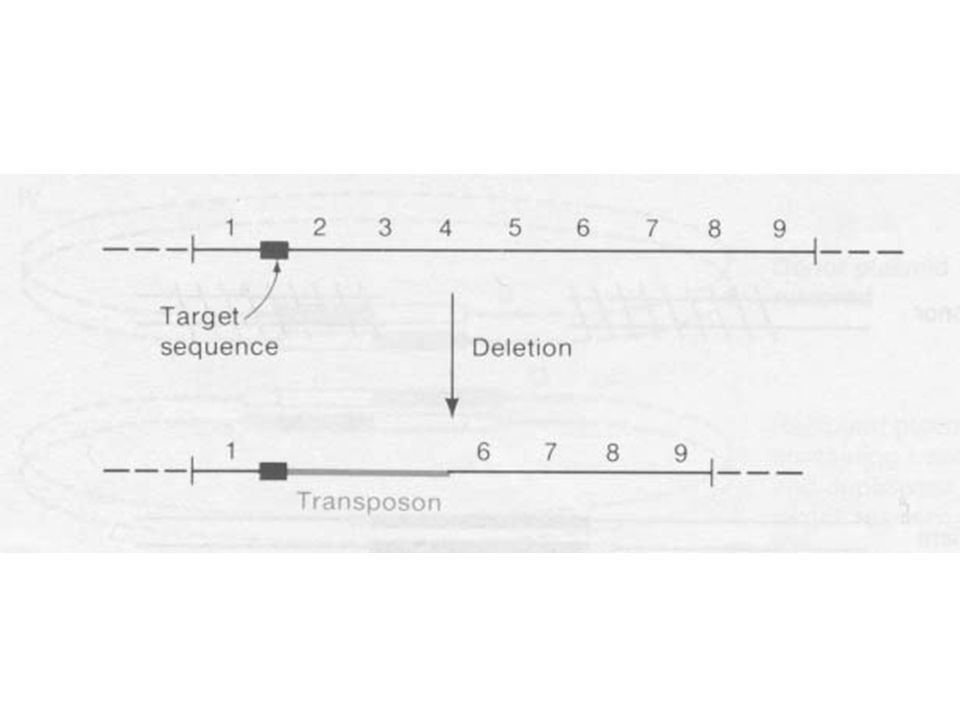
A transzpozonok hatása a baktérium genomra
A transzpozonok számos genetikai jelenséget okoznak, gén átrendeződés (rearrangement), plazmid integrációt kromoszómába Deléció A transzpozonok mellett a deléció gyakorisága x-es A deléció az elem egyik végétől indul A transzpozont egy target szekvencia határolja csak
, plazmid integrációt kromoszómába. Deléció. A transzpozonok mellett a deléció gyakorisága x-es. A deléció az elem egyik végétől indul. A transzpozont egy target szekvencia határolja csak.")
55
Mindkettőt magyarázza a transzpozíciós modell
Inverzió A transzpozon melletti inverziók gyakorisága is nagyobb Mindkettőt magyarázza a transzpozíciós modell

58
Tn5 konzervatív cut and paste

59
Direkt ismétlődés a beépüléskor

60
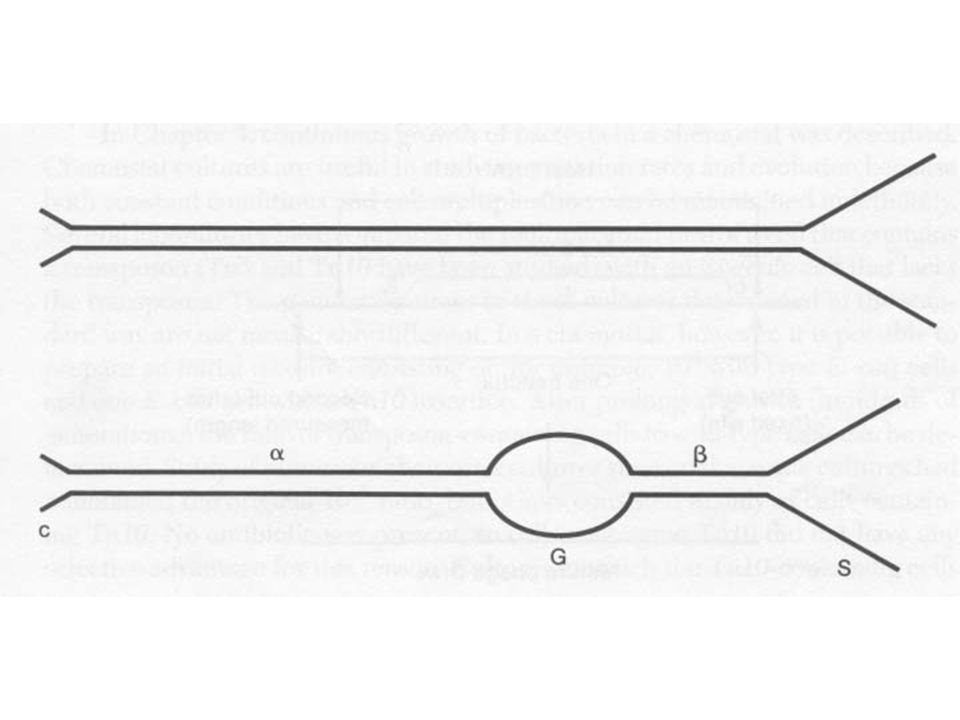
Mu fág A Mu mérsékelt fág (mutátor)
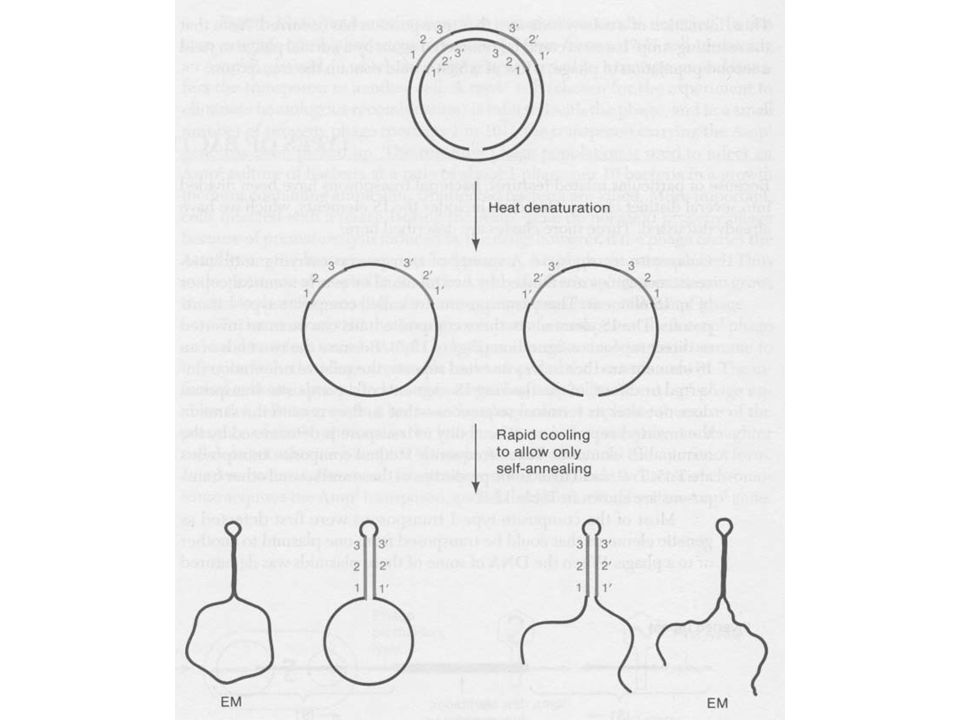
38 kb DNS, melynek végei különbözők Mu fág DNS denaturáció és renaturáció után komplementer és nem komplementer részeket mutat A nem komplementer szekvencia baktérium eredetű, minden fág részecskében más Ha a Mu DNS alfa régiójának hossza változik, az egyik nem komplementer vég hossza állandó, míg a másik (hosszabb) változik G régió, inverzió lehet, a gazdaspecifikusságért felelős
 változik. G régió, inverzió lehet, a gazdaspecifikusságért felelős.")
62
A Mu DNS replikációja és érése
Mu fág fertőzéskor mindig bekövetkezik a transzpozíció Az inzerció során csak a fág DNS épül be (a baktérium eredetű részek nem) A Mu DNS replikációja együtt jár a Mu fág beépülésével a baktérium kromoszóma különböző helyeire Ez utóbbi miatt rendelkezik különböző bakteriális szekvenciákkal
 A Mu DNS replikációja együtt jár a Mu fág beépülésével a baktérium kromoszóma különböző helyeire. Ez utóbbi miatt rendelkezik különböző bakteriális szekvenciákkal.")
64
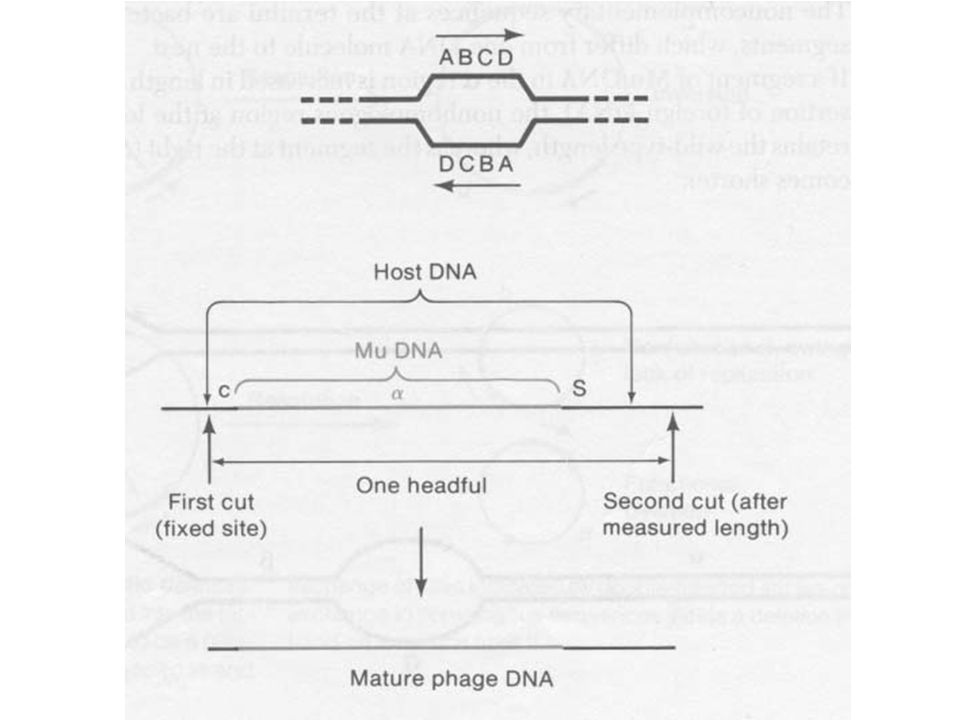
A Mu DNS mindig tartalmaz idegen darabot
A fág érése során a pakoló rendszer felismeri a Mu fágot (integrált) Hasítja a DNS-t 100 bp-al a Mu fág c vége felől Ezután fejméretnyi DNS-t pakol. Ha kell a Mu fág DNS-en kívül baktérium DNS is pakolódik
 Hasítja a DNS-t 100 bp-al a Mu fág c vége felől. Ezután fejméretnyi DNS-t pakol. Ha kell a Mu fág DNS-en kívül baktérium DNS is pakolódik.")
65
Mu fág replikatív transzpozíció

66
A Mu integráció véletlenszerű
A beépülés valószínűsége egy génbe arányos a gén nagyságával Mivel a transzpozonok sok helyre épülhetnek be és erős poláris hatást biztosítanak, továbbá az inzerció követhető (antibiotikum rezisztencia) mutáns törzsek előállítására alkalmasak Törzs fejlesztés például
 mutáns törzsek előállítására alkalmasak. Törzs fejlesztés például.")
67
Tn7 helyspecifikus transzpozíció

68
Retrotranszpozíció

69
Konjugatív transzpozíció

70
A transzpozonok és az evolúció
Antibiotikum rezisztencián kívül nincs hatása Miért marad fent evolúciós léptékben „Selfish” azaz önző gén Ha a transzpozíció gyakorisága nagyobb, mint a mutációs ráta, vagy az elem elvesztésének gyakorisága Ez elegendő a fennmaradáshoz Kemosztát kultúrában vizsgálták Kiindulás 105 baktérium, egy Tn Eredmény néhol ugyanez az arány Néhol elterjedt a Tn, mert nagyobb mértékben (jótékony mutáció) szaporodott a Tn-t tartalmazó sejt
 szaporodott a Tn-t tartalmazó sejt.")
71
A replikatív transzpozíció miatt elvileg elszaporodhatnának
Transzpozíció sokszor letális Egészséges arány kell legyen Ezt pontos szabályozás hajtja végre, nem korlátlan működés Ahol nincs transzpozon, a sejtben ott hamar elszaporodik Egy idő után stabilizálódik az elemek száma Prokariótákban általában 10-nél kevesebb van Eukariótákban a DNS 30%-a is tartalmazhat transzpozont az elemek száma is lehet

72
Transzpozonok? T-DNS (Ti plazmid) Patogenitás szigetek
Más genomi szigetek Széleken ismétlődő szekvenciák

73
Innen oda

74
Innen oda

75
Innen oda

76
Innen oda

78
IS91

79
IS91 mechanizmus

80
IS chirality

81
IS kointegrátum

84
Tc mariner

85
Insertion Sequences http://www-is.biotoul.fr/is.html
MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, Sept. 1998, p. 725–774 Vol. 62, No. 3 Insertion Sequences JACQUES MAHILLON AND MICHAEL CHANDLER Modern Microbial Genetics 17. fejezet

Hasonló előadás





 DNS-ből,>")











 Bihari Péter.>")






