Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
A természetes immunitás alapjai
2013. szeptember 18. © Dr. Fülöp A. Kristóf PhD egyetemi docens Genetikai, Sejt és Immunbiológiai Intézet Semmelweis Egyetem

2
Az immunválasz Szerzett / adaptív Természetes / veleszületett

3
A veleszületett és az adaptív immunitás összehasonlítása
Latencia nincs van Specificitás korlátozott nagy Erősítés lineáris exponenciális Memória Antigénnel való kölcsönhatás random szelektív Evolúció ősibb és konzervatívabb újabb Genetikai variábilitás sejtjei genetikailag azonosak sejtes klonalitás

4
A természetes és adaptív iv. egymásra épül és összefonódik
Magyar Tudomány, 2004/10 4

5
Az immunválasz időzítése
A természetes és adaptív immunválasz funkcionális kapcsolata emlősökben Magyar Tudomány, 2004/ o. The role of innate immunity in the organism is the identi.cation of foreign substances and their removal by phagocytosis or nitric oxide (NO) production and the recruitment of immune cells to sites of infection or in.ammation by producing cytokines. Thereby, activation of the complement cascade and the adaptive immune system by antigen presentation results in the in.ammatory response. We summarize here what is currently known about PRRs and the possible ligands to shedmore light on the pathogenesis of RA and on possible targets for novel therapies. 5 5
 production and. the recruitment of immune cells to sites of infection or in.ammation. by producing cytokines. Thereby, activation of the complement. cascade and the adaptive immune system by antigen presentation. results in the in.ammatory response. We summarize here what is. currently known about PRRs and the possible ligands to shedmore. light on the pathogenesis of RA and on possible targets for novel. therapies")
6
A természetes immunrendszer
Barrierek Makromolekulák Sejtek

7
Barrierek

8
A természetes immunrendszer szolubilis molekuláris rendszerei
Komplement rendszer Szerin proteázok rendszere, mely inaktív formában van jelen a testfolyadékokban. Aktívlótás után kaszkádreakciójuk indul be. Egyéb antimikrobiális molekulák defenzinek lizozim laktoperoxidáz laktoferrin

9
Defenzinek ++++ hydrophobic Legtöbbjük amphipathicus, töltött (rózsaszín) és hidrofób (zöld) aminosav oldalláncokkal. Pórusokat hoznak létre a sejtmembránon.
 és hidrofób (zöld) aminosav oldalláncokkal. Pórusokat hoznak létre a sejtmembránon.")
10
(gingival crevicular fluid)
Emelkedett defenzin szintek krónikus gyulladás (pl. psoriasis) colorectalis rák Defenzin deficiencia Crohn betegség?
 colorectalis rák. Defenzin deficiencia. Crohn betegség")
11
Lizozim (peptidoglycan N-acetylmuramoylhydrolase)
Könnyben, a nyálban, a verejtékben, az anyatejben, az orrváladékban és a gyomornedvben, neutrofilekben, egyes növényekben (pl. torma) is előfordul. Reduced lysozyme levels have been associated with bronchopulmonary dysplasia in newborns. Since lysozyme is a natural form of protection from gram-positive pathogens like Bacillus and Streptococcus In certain cancers (especially myelomonocytic leukemia) excessive production of lysozyme by cancer cells can lead to toxic levels of lysozyme in the blood. High lysozyme blood levels can lead to kidney failure. Source: mucosal glands Lizozim deficiencia újszülöttkori bronchopulmonary dysplasia diarrhea (tápszer) Lizozim-klorid tabletta
 is előfordul. Reduced lysozyme levels have been associated with bronchopulmonary dysplasia in newborns. Since lysozyme is a natural form of protection from gram-positive pathogens like Bacillus and Streptococcus. In certain cancers (especially myelomonocytic leukemia) excessive production of lysozyme by cancer cells can lead to toxic levels of lysozyme in the blood. High lysozyme blood levels can lead to kidney failure. Source: mucosal glands. Lizozim deficiencia. újszülöttkori. bronchopulmonary dysplasia. diarrhea (tápszer) Lizozim-klorid tabletta.")
12
Lakto- Könnyben, a nyálban, a verejtékben, az anyatejben, orrváladékban antimikrobiális hatásúak. Laktoperoxidáz rendszer acceptor + H2O2 → oxidized acceptor + H2O hatásos általában aerob mikroorganizmusok ellen laktoferrinnel és lizozimmel szinergizál Termelik: neutrofilek és mucosalis mirigyek Fogkrémekbe is adagolhatják Laktoferrin Hatás vas kötés peroxidáció Termelik: neutrofilek és mucosalis mirigyek Source: mucosal glands lactoperoxidase system does not attack DNA and is not mutagenic. Source of the hydrogen peroxide (H2O2) usually is the reaction of glucose with oxygen in the presence of the enzyme glucose oxidase (EC ) that also takes place in saliva.
 usually is the reaction of glucose with oxygen in the presence of the enzyme glucose oxidase (EC ) that also takes place in saliva.")
13
A természetes immunválasz sejtjei
Monocita – makrofág rdsz. Neutrofil Hízósejt Dendritikus sejtek (DC) Innate lymphoid cells (ILCs) pl. NK (natural killer) Natural lymphocytes: B1 B cells, NKT cells and T cells
 Innate lymphoid cells (ILCs) pl. NK (natural killer) Natural lymphocytes: B1 B cells, NKT cells and T cells.")
14
Innate lymphoid cells (ILCs)
Csoportosításuk a citokin termelésen alapszik ILCs1 csoport I: IFN gamma (NK cell) anti-tumor és anti-virális védekezés a limfoid szövetek fejlődése (embriogenezis) ILCs2 : Th2 citokinek (pl. IL5, 13) Szöveti regeneráció, parazita ellenes hatás, allergia, asztma ILCs3: IL17 vagy IL22 Egyensúly a gasztrointesztinális szimbionták és az immunválasz között, inflammatory bowel diseases (IBD). Védelem az extracelluláris kórokozók ellen.
 anti-tumor és anti-virális védekezés. a limfoid szövetek fejlődése (embriogenezis) ILCs2 : Th2 citokinek (pl. IL5, 13) Szöveti regeneráció, parazita ellenes hatás, allergia, asztma. ILCs3: IL17 vagy IL22. Egyensúly a gasztrointesztinális szimbionták és az immunválasz között, inflammatory bowel diseases (IBD). Védelem az extracelluláris kórokozók ellen.")
15
A felismerés stratégiái
Nem saját PAMP (pathogen associated molecular pattern) Hiányzó saját NK: hiányzó MHC Megválto-zott saját ECM degradációs termékek, tumorantigén
 Hiányzó saját. NK: hiányzó MHC. Megválto-zott saját. ECM degradációs termékek, tumorantigén.")
16
NK (natural killer / természetes ölő) sejtek
Nem T, nem B limfociták Nagy citolplazmatikus granulumokkal (LGL: Large electrondense garnules) Tumor és vírusfertőzött sejtek elpusztítása 10-15%-a a prifériás limfocitáknak
 Tumor és vírusfertőzött sejtek elpusztítása %-a a prifériás limfocitáknak.")
17
Az NK sejtek morfológiája
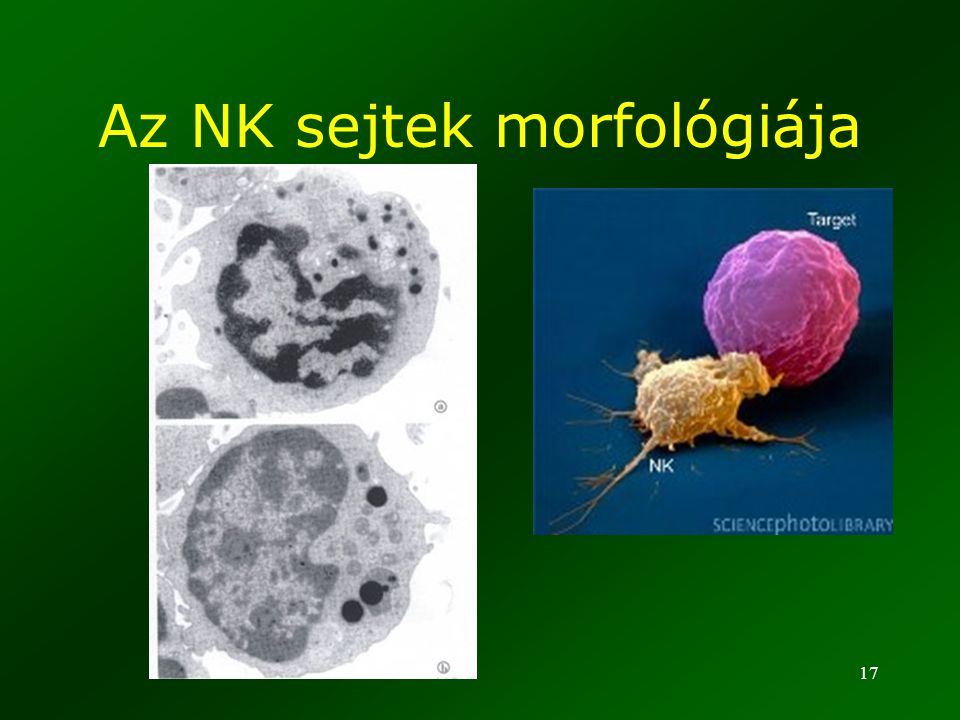
18
(KAR: MICA (sterssz indukált glikoprotein)
(KAR: MICA (sterssz indukált glikoprotein) KIR: pl. HLA-E R HLA-C R FC gamma R KIR: Killer inhibitory receptor, KAR: killer activating receptor
 KIR: pl. HLA-E R. HLA-C R. FC gamma R. KIR: Killer inhibitory receptor, KAR: killer activating receptor.")
19
Mit ismer fel az NK sejt?

20
Az Nk sejt effektor válasza

21
Az NK (és TCT) sejt effektor molekulái
Perforin degarnuláció után a célsejt membránját kilyukasztja strukturálisan és funkcionálisan hasonló C9-hez Granzimek (granulum enzimek) szerin proteázok a perforin által formált póruson (vagy M6P receptorral) bejut a célsejtbe és kaszpáz aktiváció útján apoptózist indukál
 szerin proteázok. a perforin által formált póruson (vagy M6P receptorral) bejut a célsejtbe és. kaszpáz aktiváció útján. apoptózist indukál.")
22
A felismerés stratégiái
Nem saját PAMP (pathogen associated molecular pattern) Hiányzó saját NK: hiányzó MHC Megválto-zott saját ECM degradációs termékek, tumorantigén
 Hiányzó saját. NK: hiányzó MHC. Megválto-zott saját. ECM degradációs termékek, tumorantigén.")
23
Dendritikus sejtek (DC)
Fagocita aktivitás Nincs fagocita aktivitás Antigén prezentálás

24
A DC család Mieloid DC CD141+ CD11c+ Monocyte derived DC (MDDC)
Monocitához hasonló IL-12, TLR3 MHC I (kereszt) prezentáció, CD8+ T, Th1 CD11c+ (pl. Langerhans sejtek) Kemokinek MHC II prezentáció CD4+ T Monocyte derived DC (MDDC) Monocitából in vitro Plazmacitoid DC Plazma sejthez hasonló INF alfa, TLR 7,9 memória Follicular DC Mesenchimális, nincs MHCII. Natív antigén prezentálás a germinális centrumokban.
 prezentáció, CD8+ T, Th1. CD11c+ (pl. Langerhans sejtek) Kemokinek. MHC II prezentáció CD4+ T. Monocyte derived DC (MDDC) Monocitából in vitro. Plazmacitoid DC. Plazma sejthez hasonló. INF alfa, TLR 7,9. memória. Follicular DC. Mesenchimális, nincs MHCII. Natív antigén prezentálás a germinális centrumokban.")
25
A DC érése

26
DC, az immunrendszer „karmestere”

27
DC, az immunrendszer „karmestere” (2)
")
28
Fagocitózis Elie Metchnikoff (1845–1916) Chemotaxis Abszorpció
Receptorok Mannóz-R LPS-R Opszonizáció

29
FC receptor mediált fagocitózis
pseudopodium 29

30
Endoszomális lizoszómális útvonal
phagocytosis receptor endocytosis receptor Actin polymerization Clathrin-coated pit phagosome endosomes Fusion with endosomes and lysosomes Gogi Hydrolytic enzymes lysosomes Phagolysosome 30

31
Frusztrált fagocitózis
A fagocita a nagy patogén (parazita) felszínéhez kötődik A lizoszómális tartalom exocitózissal ürül a extracelluláris térbe 31
 felszínéhez kötődik. A lizoszómális tartalom exocitózissal ürül a extracelluláris térbe. 31.")
32
Frusztrált fagocitózis (2)
")
33
MPS (mononuclearis fagocita rdsz.)
Szövetspecifikus makrofágok

34
Makrofág differenciálódás és aktiváció

35
Makrofág altípusok aktiváció szerint
FIZZ1 is a novel macrophage gene in chronic Th2-mediated inflammation.

36
A makrofágok bekebelezik és lebontják a megkötött baktériumokat
A makrofágon többféle receptor különféle bakteriális részeket ismer fel 36 36

37
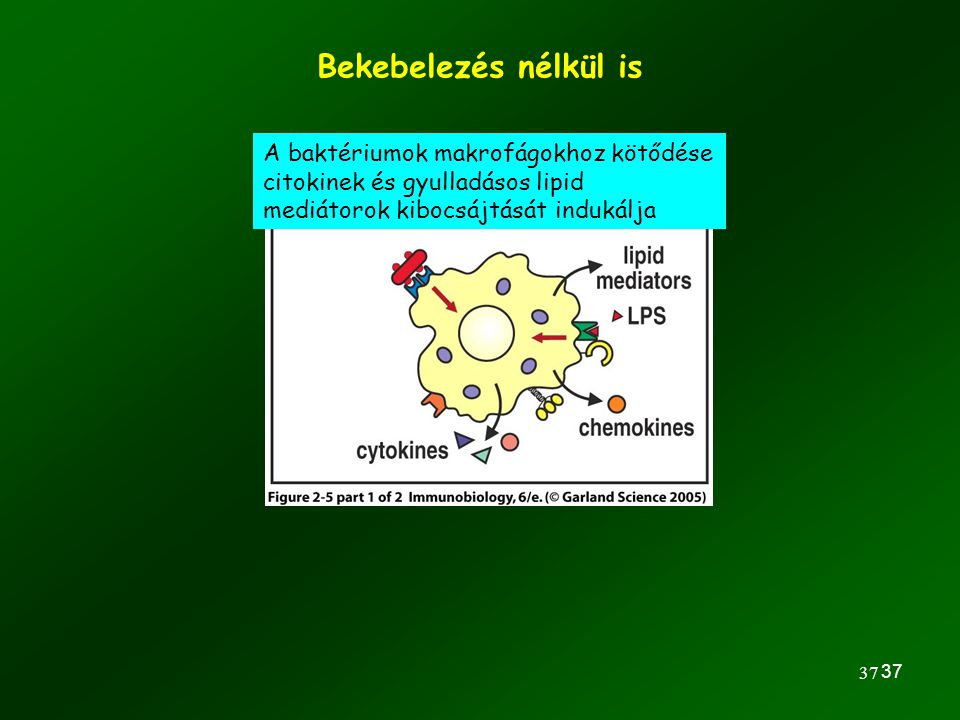
Bekebelezés nélkül is A baktériumok makrofágokhoz kötődése citokinek és gyulladásos lipid mediátorok kibocsájtását indukálja 37 37
38
Neutrophil granulocita differenciálódása és aktivációja

39
Neurofilek antimikrobiális hatása
Extracelluláris csapda Nature Reviews Immunology 9, (2009) intravezikuláris ölés (oxigén függő és független)
 intravezikuláris ölés. (oxigén függő és független)")
40
NADPH oxidase

41
The growth of Staphylococcus aureus is potently inhibited by NO, Salmonella typhimurium, Escherichia coli and Listeria monocytogenes seem to be resistant.

42
A fagociták meg tudják különböztetni a sejthalálokat
Necrosis

43
Az apototikus sejtek / testek fagocitózisa nem indukál gyulladást
Apoptotikus sejt Plazma membrán: sziálsav foszfatidil szerin externalizáció ”eat me „ szignál fagocita Nincs gyulladásos citokin termelés

44
A fagociták szerepe többrétű
Antigének felvétele – antigén prezentálás Eliminálás patogének apoptotikus sejtek nekrotikus sejttörmelék ECM dagradációs termékek Szabályozó anyagok termelése pl. gyulladásos citokinek

45
Fagocita deficienciák (számos típus)
Pl. Krónikus granulomatozus betegség (CGD) Kevesebb és kevésbé aktív fagociták NADH/NADPH oxidáz mutáció Csökkent oxigén függő baktérium-ölés INF gamma kezelés fogágybetegség
 Kevesebb és kevésbé aktív fagociták. NADH/NADPH oxidáz mutáció. Csökkent oxigén függő baktérium-ölés. INF gamma kezelés. fogágybetegség.")
46
Fagocita deficienciák II.
Leukocita adhéziós deficiencia Magas vér neu. Csontvelő transzplantáció Fagocita deficienciák (folyt.) Chediak-Higashi szindróma lizoszomális targeting neutrofil deficiencia parodontitis fogágybetegség
 Chediak-Higashi szindróma. lizoszomális targeting. neutrofil deficiencia. parodontitis. fogágybetegség.")
47
A fagociták receptorai
Opszonizáló receptorok Fc Komplement Lektin R Opszonizáció: tálalás a fagociták számára Mintázatfelismerő(PRR-Pattern Recognition Receptors)
")
48
Opszonizáció baktérium Fagocita Lektin receptor Fc receptor
Komplement receptor Fc receptor Fagocita szénhidrát Lektin receptor Antigén-antitest Komplement komponens

49
Immunglobulin Fc régió
papain Fc region Fc= fragment crystallizable 49

50
Fc receptorok Fcg receptors Fce receptors IgG PolyIg receptor
opszonizáció (e.g. FcRI) respiratory burst B-sejt szabályozás (FcR IIb) Fce receptors IgE nagy affinitású (FcRI) hízósejteken és bazifileken (allergia) alacsony affinitásaú (CD23): szabályozás polimer (IgA) transzcitózis (mucosa) PolyIg receptor FcRn receptor (neonatal) IgG transzcitózis újszülött passzív immmunizálása degradáció elleni védelem 50
 respiratory burst. B-sejt szabályozás (FcR IIb) Fce receptors. IgE. nagy affinitású (FcRI) hízósejteken és bazifileken (allergia) alacsony affinitásaú (CD23): szabályozás. polimer (IgA) transzcitózis (mucosa) PolyIg receptor. FcRn receptor (neonatal) IgG. transzcitózis. újszülött passzív immmunizálása. degradáció elleni védelem. 50.")
51
Poli-Ig receptor és transzcitózis

52
FcRn (neonatal) kettős funkciója
 kettős funkciója")
53
Mintázatfelismerő receptorok
Membrane PRR (pl.TLR) Szecernált PRR Intracelluláris PRR cytosolic vesicular 53
 Szecernált PRR. Intracelluláris PRR. cytosolic. vesicular. 53.")
54
Mintázat felismerő molekulák
Extracelluláris Pentraxinok (pl. CRP) Plazma membrán Toll like receptorok (pl. TLR4) Vezikuláris pl. TLR3 Citoszol NOD like receptorok (Pl. NOD-1)
 Plazma membrán. Toll like receptorok (pl. TLR4) Vezikuláris. pl. TLR3. Citoszol. NOD like receptorok (Pl. NOD-1)")
55
Felismerés - több ponton
Szecernált PRR opszonizálás Endoszomális PRR Nat Rev Immunol Jan;6(1):33-43.
:")
57
TOLL-LIKE RECEPTOROK (TLR)
horog alak leucin-gazdak ismétlődő szekvenciák PAMP (pathogen associated molecular patterns) felismerés evolúciósan konzerváltak 57
 felismerés. evolúciósan konzerváltak. 57.")
58
TLR-ek ligandjai (1)
")
59
TLR-ek ligandjai (2) A sejtfelszínen elhelyezkedő TLR-ek olyan ligandokat ismernek fel, amilyen egyáltalán nincs a gazdaszervezetben: pl. LPS, flagellin Az intracellulárisan elhelyezkedő TLR-ek nukleinsav alapú PAMP-ot ismernek fel.

60
A TLR-ek családfája

61
A PRR-ek szabják meg, hogy milyen gyulladásos és immunmodulátor citkinek termelődnek

62
Inflammasome inducers
A szignalizációban rész vevő nagy intracitoplazmatikus komplexeket inflammaszómáknak nevezik Inflammasome inducers Alternatív szekréció

63
TLR gátlás a terápiában (krónikus gyulladás)
TLR antagonisták endoszómális TLR-DNS interakciót gátló szerek Pl. chloroquin erős bázis, ami az endoszóma savasodását gátolja As it mildly suppresses the immune system, it is used in some autoimmune disorders, such as rheumatoid arthritis and lupus erythematosus. Hans-Joachim Anders, 1 Daniel Zecher,1 Rahul D Pawar,1 and Prashant S Patole1 Molecular mechanisms of autoimmunity triggered by microbial infection Arthritis Res Ther. 2005; 7(5): 215–224. suppressive ODN may represent a functional antagonist for TLR9 signalling induced by CpGDNA, a hypothesis supported by our studies with MRLlpr/lpr mice. These findings support a role for TLR9 signalling in the pathogenesis of lupus. Thus, developing specific small molecule TLR9 antagonists may represent a new approach as a preventive therapy for systemic lupus erythematosus. Gyógyszeres malária prevenció heti 2 tabl. Chloroquin (Delagil) adása 2008: Small-molecule TLR9 inhibitors are being developed (Sally Ishizaka, Andover, Massachusetts, USA) and the nucleic acid–based modulators of the endosomal TLRs are progressing to clinical testing (Sudhir Agrawal, Cambridge, Massachusetts, USA, and Robert Coffmann, Berkeley, California, USA). TLR-szignalizációt gátló szerek csökkentik a gyulladásos citokineket kódoló gének expresszióját 63
: 215–224. suppressive ODN may represent a. functional antagonist for TLR9 signalling induced by CpGDNA, a hypothesis supported by our studies with MRLlpr/lpr. mice. These findings support a role for TLR9 signalling in the. pathogenesis of lupus. Thus, developing specific small. molecule TLR9 antagonists may represent a new approach. as a preventive therapy for systemic lupus erythematosus. Gyógyszeres malária prevenció heti 2 tabl. Chloroquin (Delagil) adása. 2008: Small-molecule. TLR9 inhibitors are being developed (Sally. Ishizaka, Andover, Massachusetts, USA) and the nucleic acid–based modulators. of the endosomal TLRs are progressing to. clinical testing (Sudhir Agrawal, Cambridge, Massachusetts, USA, and Robert Coffmann, Berkeley, California, USA). TLR-szignalizációt gátló szerek csökkentik a gyulladásos citokineket kódoló gének expresszióját. 63.")
64
Mi a helyzet a kommenzális flórával?

65
TLR-ek nem expresszálódnak az apikális felszínen

66
Pattern associated molecular pattern DAMP:
PAMP: Pattern associated molecular pattern DAMP: Damage asssociated molecular pattern Dendritikus sejt Kostimulációs molekula Nature Reviews Immunology 8, (April 2008) Nature Reviews | Immunology Stranger and danger models. Dendritic cells (DCs) are present in all tissues, where they gather antigens from the local environment but are not in an immunostimulatory state. In Janeway’s ‘stranger’ model3, antigen-presenting cells (later appreciated to be DCs) were endowed with pattern-recognition receptors (PRRs) that recognize the unique features of microbial molecules (pathogen-associated molecular patterns, PAMPs). When PAMPs were present — for example, from an infection or adjuvant — then DCs were stimulated to migrate to lymphoid tissues and present both antigen and co-stimulatory molecules (CD80 and/or CD86) to T cells. In Matzinger’s ‘danger’ model7, the crucial event controlling the initiation of an immune response was not infection, but the production of danger signals known as damage-associated molecular patterns (DAMPs) from cells stressed, damaged and/or dying in the local tissue. These were postulated to act on DCs in a manner that also caused them to migrate to lymphoid tissue and present antigens to T cells in an immunostimulatory manner. It has been speculated that DAMPs might be produced in response to PAMPs and therefore that DAMPs might be the final mediator promoting immune responses in all situations, including infection. This might occur; however, it is also possible, and in our view probable, that DAMPs and PAMPs can alert the immune system to a problem independently and possibly even in a synergistic manner. TCR, T-cell receptor. dying cells were postulated to release endogenous adjuvants that, using similar nomenclature to PAMPs, have been called damage-associated molecular patterns (DAMPs); dead cells contain endogenous adjuvants that promote T-cell immunity. It was also found that dead cells could stimulate dendritic cells (DCs) to mature into immunostimulatory cells in culture10 and could promote the migration of mature DCs into draining lymph nodes in animals12. Mi a DAMP receptor? several purified DAMPs have been reported to stimulate TLR2 and TLR4. In addition to TLRs, some other receptors have been implicated in the actions of various DAMPs (TABLeS 1,2). RAGe (receptor for advanced glycation end-products; also known as AGeR) binds HMGB1 and s100 proteins Galectins have been reported to stimulate glycosylated surface receptors such as CD2 (ReF. 40). Integrins90, chemokine receptors69 and CD44 (ReF. 91) have been reported to be the targets of DAMPs from extracellular-matrix components. . Vándorlás a nyirokcsomóba 66
 Nature Reviews | Immunology. Stranger and danger models. Dendritic cells (DCs) are present in all. tissues, where they gather antigens from the local environment but are not in an. immunostimulatory state. In Janeway’s ‘stranger’ model3, antigen-presenting cells (later. appreciated to be DCs) were endowed with pattern-recognition receptors (PRRs) that. recognize the unique features of microbial molecules (pathogen-associated molecular. patterns, PAMPs). When PAMPs were present — for example, from an infection or. adjuvant — then DCs were stimulated to migrate to lymphoid tissues and present both. antigen and co-stimulatory molecules (CD80 and/or CD86) to T cells. In Matzinger’s. ‘danger’ model7, the crucial event controlling the initiation of an immune response was. not infection, but the production of danger signals known as damage-associated. molecular patterns (DAMPs) from cells stressed, damaged and/or dying in the local. tissue. These were postulated to act on DCs in a manner that also caused them to migrate. to lymphoid tissue and present antigens to T cells in an immunostimulatory manner. It has. been speculated that DAMPs might be produced in response to PAMPs and therefore. that DAMPs might be the final mediator promoting immune responses in all situations, including infection. This might occur; however, it is also possible, and in our view. probable, that DAMPs and PAMPs can alert the immune system to a problem. independently and possibly even in a synergistic manner. TCR, T-cell receptor. dying cells were postulated to release endogenous. adjuvants that, using similar nomenclature to PAMPs, have been called damage-associated molecular patterns. (DAMPs); dead cells contain. endogenous adjuvants that promote T-cell immunity. It. was also found that dead cells could stimulate dendritic. cells (DCs) to mature into immunostimulatory cells in. culture10 and could promote the migration of mature. DCs into draining lymph nodes in animals12. Mi a DAMP receptor several purified DAMPs have been reported to. stimulate TLR2 and TLR4. In addition to TLRs, some other receptors have been. implicated in the actions of various DAMPs (TABLeS 1,2). RAGe (receptor for advanced glycation end-products; also known as AGeR) binds HMGB1 and s100 proteins. Galectins have been reported to stimulate. glycosylated surface receptors such as CD2 (ReF. 40). Integrins90, chemokine receptors69 and CD44 (ReF. 91) have been reported to be the targets of DAMPs from. extracellular-matrix components. . Vándorlás. a nyirokcsomóba. 66.")
67
Veszély csak nekrotikus sejthalál vagy ECM degradáció esetén
DAMP - Hősokk fehérjék - HMGB1 : intracelluláris DNS kötő fehérje - genomiális dupla szálú DNS extracelluláris molekulák: pl. fragmentált kollagén, hialuronsav … Nature Reviews Immunology 8, (April 2008) most DAMPs come from pre-existing molecules then how does the innate immune system distinguish dead cells from live ones? The crucial event upon necrotic cell death is thought to be the release of DAMPs from intracellular stores, which we refer to as the hidden self model when cells undergo necrosis they lose the integrity of their plasma membrane and release their intracellular contents, including the cytosolic DAMPs, into the extracellular milieu. This can also occur when apoptotic cells aren’t rapidly cleared Discriminating between viable cells, necrosis and apoptosis. The hidden self model proposes that innate immune cells have receptors that detect certain intracellular molecules, known as damageassociated molecular patterns (DAMPs), that are normally hidden in the interior of cells and that are only revealed after necrosis. This model can explain why live cells, which contain pre-existing danger signals, don’t stimulate the innate immune system. Moreover, it can also explain why necrotic cells always stimulate the innate immune system, whereas apoptotic cells are only stimulatory in some situations99 and not in others100. This is because necrotic cells always lose membrane integrity and release their intracellular contents. By contrast, apoptotic cells initially maintain membrane integrity. If apoptotic cells are then rapidly cleared by phagocytes, the dead cells don’t release their intracellular DAMPs and the immune system is not stimulated. However, if apoptotic cells are not rapidly cleared, as might occur in a solid organ, they undergo secondary necrosis and become permeable. The resulting release of DAMPs then stimulates the innate immune system. So, the event that communicates ‘danger’ to the immune system is not how the cell dies per se, but whether or not it eventually loses membrane integrity and releases its intracellular contents into the extracellular milieu. This concept of hidden signals being revealed might also apply to DAMPs of extracellular origin (for example, when revealed through the action of enzymes released after cell damage) and even, as has been proposed, hidden portions of molecules8. Nincs válasz, vagy anti-inflammatorikus válasz Pro-inflammatorikus válasz 67
 most DAMPs come from pre-existing molecules then how does. the innate immune system distinguish dead cells from. live ones The crucial event upon necrotic cell death is. thought to be the release of DAMPs from intracellular. stores, which we refer to as the hidden self model. when cells undergo necrosis they lose. the integrity of their plasma membrane and release. their intracellular contents, including the cytosolic. DAMPs, into the extracellular milieu. This can also. occur when apoptotic cells aren’t rapidly cleared. Discriminating between viable cells, necrosis and apoptosis. The hidden self model proposes. that innate immune cells have receptors that detect. certain intracellular molecules, known as damageassociated. molecular patterns (DAMPs), that are normally. hidden in the interior of cells and that are only revealed. after necrosis. This model can explain why live cells, which. contain pre-existing danger signals, don’t stimulate the. innate immune system. Moreover, it can also explain why. necrotic cells always stimulate the innate immune system, whereas apoptotic cells are only stimulatory in some. situations99 and not in others100. This is because necrotic. cells always lose membrane integrity and release their. intracellular contents. By contrast, apoptotic cells initially. maintain membrane integrity. If apoptotic cells are then. rapidly cleared by phagocytes, the dead cells don’t release. their intracellular DAMPs and the immune system is not. stimulated. However, if apoptotic cells are not rapidly. cleared, as might occur in a solid organ, they undergo. secondary necrosis and become permeable. The resulting. release of DAMPs then stimulates the innate immune. system. So, the event that communicates ‘danger’ to the. immune system is not how the cell dies per se, but whether. or not it eventually loses membrane integrity and releases. its intracellular contents into the extracellular milieu. This. concept of hidden signals being revealed might also apply. to DAMPs of extracellular origin (for example, when. revealed through the action of enzymes released after cell. damage) and even, as has been proposed, hidden portions. of molecules8. Nincs válasz, vagy anti-inflammatorikus válasz. Pro-inflammatorikus válasz. 67.")
68
Az immunválasz aktiválódásához több szignál kell
Mintázat felismerő receptor Kostimuláció: APC - Th CD80/B7-CD28 Th - B CD40L-CD40 Veszély szignál Antigén

69
A természetes immunitáshoz tartozik
De más foglakozásokon kerül szóba: kemotaxis, sejtmigráció komplement gyulladás

70
V É G E

Hasonló előadás




















