Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Endoszimbionta sejtorganellumok II.
A peroxiszóma, a centriólum és a színtest. 2011. November 14. Dr.Tóth Sára Egyetemi docens Genetikai, Sejt- és Immunbiológiai Intézet

3
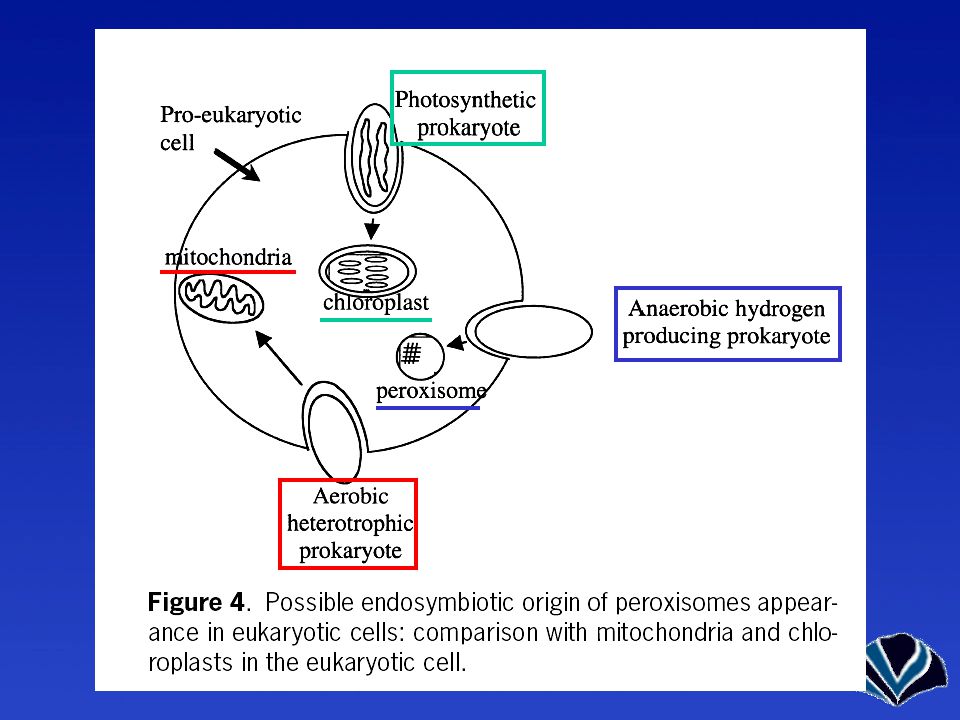
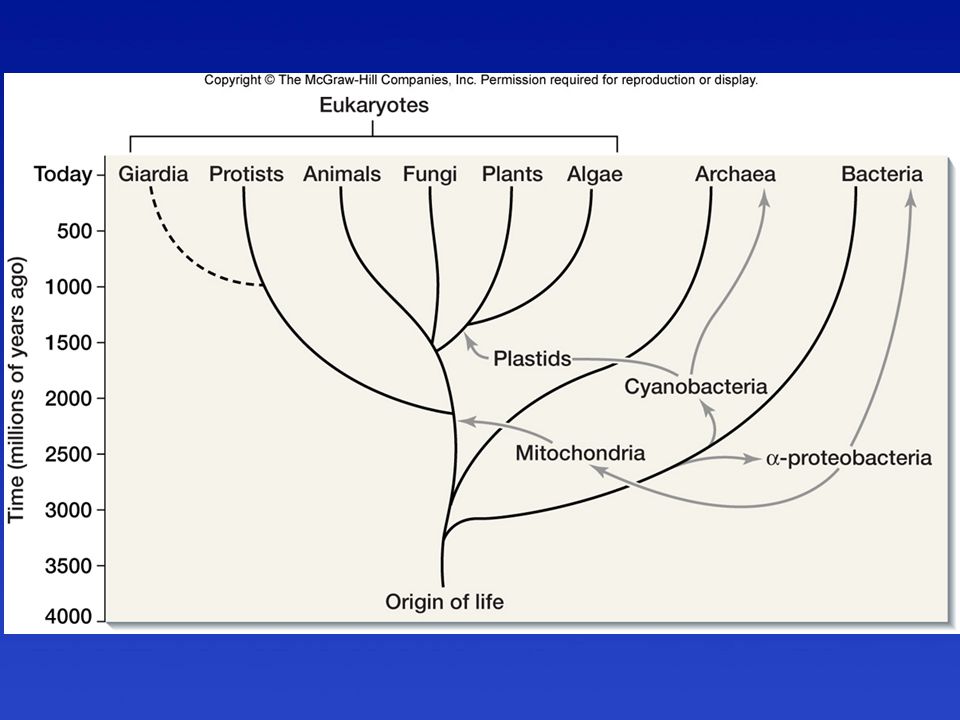
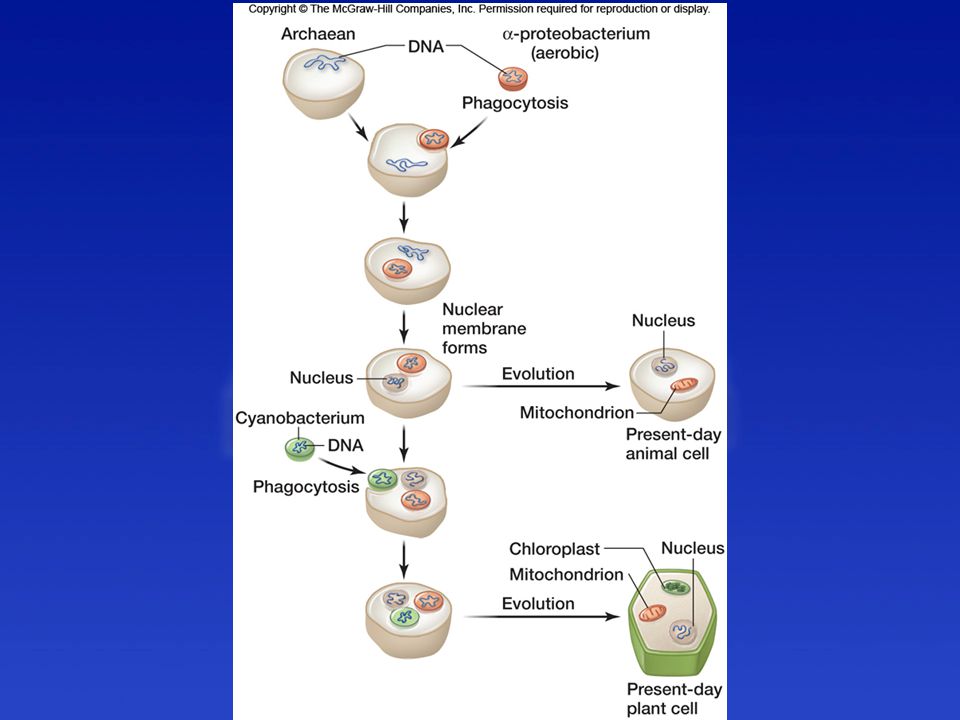
Endoszimbionták = sejtorganellumok
Membrán nélkül - Centriólum Membránnal Mitokondrium Peroxiszóma Kloroplasztisz

4
A peroxiszóma jellemzői
Egyszeres membrán (egy lipid bilayer) Szelektív fehérje import Nincs DNS-e (genomja) Jellemző enzimjei: kataláz, húgysav-oxidáz (krisztalloid)
 Szelektív fehérje import. Nincs DNS-e (genomja) Jellemző enzimjei: kataláz, húgysav-oxidáz (krisztalloid)")
5
A peroxiszóma szerkezete

6
Peroxiszóma növényi sejtben
Krisztalloid Kloroplasztisz

7
Peroxiszóma állati sejtben
Krisztalloid Mátrix

8
Különböző fajok peroxiszómái

9
Peroxiszóma állati sejtben
Immuncitokémia

10
Funkciói Számos biokémiai folyamatban: RH2 + O2 = R + H2O2
Kataláz: H2O2 + R’H2 = R’ + 2H2O (méregtelenítés májban,vesében pl. metanol, etanol, fenolok) Zsírsav oxidáció (mint a mitokondriumokban, de növényekben és gombákban csak a peroxiszómákban)
 Zsírsav oxidáció (mint a mitokondriumokban, de növényekben és gombákban csak a peroxiszómákban)")
11
A peroxiszóma funkciói
Különböző anyagok szintézise glicerolipidek plasmalogen (a myelin ~80 %-át alkotó foszfolipid) koleszterin epesavak Oxidációs folyamatok Hosszú és nagyon hosszú szénláncú zsírsavak = LFA és VLCFA oxidációja (növényekben kizárólag a peroxi- szómákban) purin metabolizmus „d” aminosavak oxidációja A fenti folyamatokban melléktermékként H2O2 keletkezik. A KATALÁZ enzim eliminálja a H2O2-t (méregtelenítés) 2 H2O2 = 2 H2O + O2
 koleszterin. epesavak. Oxidációs folyamatok. Hosszú és nagyon hosszú szénláncú zsírsavak = LFA és VLCFA oxidációja (növényekben kizárólag a peroxi- szómákban) purin metabolizmus. „d aminosavak oxidációja. A fenti folyamatokban melléktermékként H2O2. keletkezik. A KATALÁZ enzim eliminálja a H2O2-t (méregtelenítés) 2 H2O2 = 2 H2O + O2.")
12
Zsírsav oxidáció a peroxiszómában
A közepes láncú zsírsavak (10-20C) a peroxiszómákban és a mitokondriumokban is lebomolhatnak. A oxidáció itt is 4-lépéses folyamat, de a FAD és a NAD által összegyűjtött elektronok közvetlenül az O2-hez jutnak, káros H2O2 keletkezik, amely a kataláz enzim által rögtön H2O-vá és O2-é alakul át. Az energia hő formájában szabadul fel, nem tárolódik ATP formájában.
 a peroxiszómákban és a mitokondriumokban is lebomolhatnak. A oxidáció itt is 4-lépéses folyamat, de a FAD és a NAD által összegyűjtött elektronok közvetlenül az O2-hez jutnak, káros H2O2 keletkezik, amely a kataláz enzim által rögtön H2O-vá és O2-é alakul át. Az energia hő formájában szabadul fel, nem tárolódik ATP formájában.")
13
peroxiszómális folyamatok összehasonlítása
A mitokondriális és peroxiszómális folyamatok összehasonlítása ! ! !

14
Az állati sejtek peroxiszómáiban termelt acetil-CoA a citoszolba kerül, ahol a koleszterin és más metabolitok szintéziséhez használódik fel. A mitokondrium és a peroxiszóma -oxidációs enzimjei eltérő módon szerveződnek: különálló enzimek a mitokondriumokban (mint a Gram-pozitív baktériumokban) és egy komplexet alkotnak a peroxiszómákban (minta Gram-negatív baktériumokban), ahol legalább két enzim van egy polipeptid láncban.
 és egy komplexet alkotnak a peroxiszómákban (minta Gram-negatív baktériumokban), ahol legalább két enzim van egy polipeptid láncban.")
15
A mitokondrium és a peroxiszóma -oxidációs
Peroxiszómákban és Gram-negatív baktériumokban Mitokondriumokban és Gram-pozitív baktériumokban A mitokondrium és a peroxiszóma -oxidációs enzimjei eltérő módon szerveződnek

17
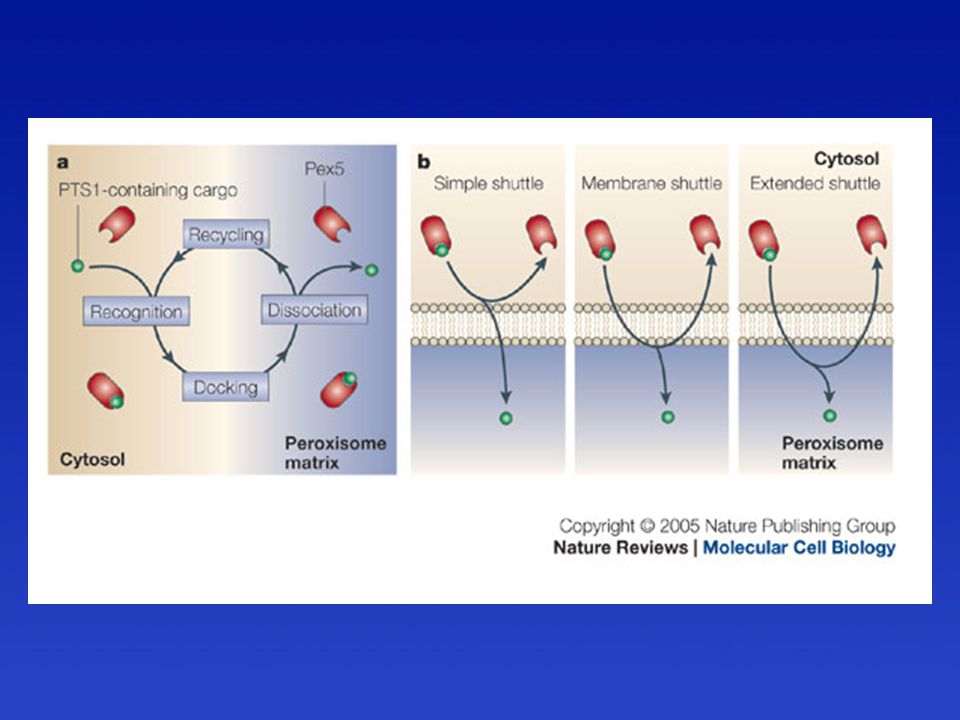
Peroxiszómális poszt-transzlációs transzmembrán transzport
A fehérje import hajtóereje az ATP hidrolízis.

18
A peroxiszóma membrán körülveszi az üres peroxiszóma mátrixot A membránon keresztül újabb fehérjék importálódnak a mátrixba Peroxiszóma membránfehérjéi beépülnek a membránba

19
Peroxiszómális szignál szekvencia PTS = Peroxisomal targeting signals
PTS1 = 3-aminosav hosszú; SKL (Ser-Lys-Leu)
")
20
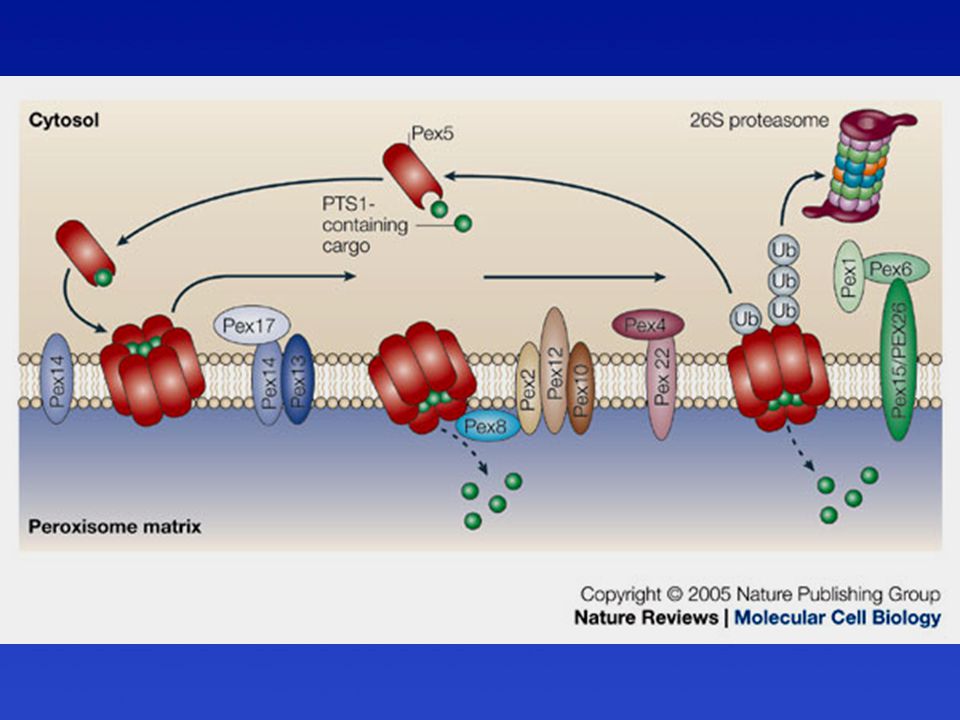
A peroxiszóma mátrix fehérjéinek transzportja
Pex-ek = peroxinok = peroxiszóma biogenezis faktorok. A peroxiszómák felszínén mint citoszol receptorok vagy dokkoló fehérjék működnek

21
Kataláz transzport a peroxiszómába
PTS1R = Pex5 Az importálandó fehérjék nem tekerednek le. Így bizonyos oligomer fehérjék egy csoportként importálódnak.

24
AAA+ proteins= ATP-ases associated with a variety of different cellular activities

25
A peroxiszóma biogenezis és osztódás modellje
PMP = peroxisomal membrane protein Alberts és mtsai: Molecular Biology of the Cell, 5. kiadás, 2008 nyomán

26
A peroxiszóma mátrix és membrán fehérjéi eltérő úton jutnak be a peroxiszómába

27
A peroxiszomális membránfehérjék A peroxiszómák osztódása:
(PMP-k) eredete A peroxiszómák osztódása: kettéhasadás Alberts és mtsai: Molecular Biology of the Cell, 5. kiadás, 2008 nyomán
 eredete. A peroxiszómák osztódása: kettéhasadás. Alberts és mtsai: Molecular Biology of the Cell, 5. kiadás, 2008 nyomán.")
28
A peroxiszómális fehérje import tisztázatlan kérdései

29
A peroxiszóma keletkezése

30
Élesztő (Yarrowia lipolytica)
Ember

31
A peroxiszóma összeszerelődés lépései (Yarrowia lipolytica)
Élesztő (Yarrowia lipolytica) Ember
 Ember.")
32
Az ER szerepe a peroxiszóma biogenezsisben
Most, if not all, PMPs are first imported into the ER through the Sec61/SSH1 translocon or the GET3 complex (left inset), are sorted into a pre-peroxisomal compartment, and bud out in a Pex3/Pex19-dependent manner to form pre-peroxisomal vesicles (right inset). These vesicles can form mature peroxisomes after fusion, dependent on Pex1/Pex6 (Titorenko and Rachubinski, 1998) and matrix protein import (de novo pathway). The de novo pathway repopulates cells with peroxisomes in the biogenesis mutants (e.g., pex3Δ/pex19Δ) lacking the organelle when corresponding genes are reintroduced (Elgersma et al., 1997; Fang et al., 2004; Tam et al., 2005; Hoepfner et al., 2005; Motley and Hettema, 2007; Motley et al., 2008; Perry et al., 2009; van der Zand et al., 2010). Alternatively, the pre-peroxisomal vesicles fuse with divided peroxisomes generated from preexisting mature peroxisomes. Peroxisome division requires Pex11 and a specific set of DRPs. In plants, retrograde trafficking from peroxisomes to the ER has been described (McCartney et al., 2005).
, are sorted into a pre-peroxisomal compartment, and bud out in a Pex3/Pex19-dependent manner to form pre-peroxisomal vesicles (right inset). These vesicles can form mature peroxisomes after fusion, dependent on Pex1/Pex6 (Titorenko and Rachubinski, 1998) and matrix protein import (de novo pathway). The de novo pathway repopulates cells with peroxisomes in the biogenesis mutants (e.g., pex3Δ/pex19Δ) lacking the organelle when corresponding genes are reintroduced (Elgersma et al., 1997; Fang et al., 2004; Tam et al., 2005; Hoepfner et al., 2005; Motley and Hettema, 2007; Motley et al., 2008; Perry et al., 2009; van der Zand et al., 2010). Alternatively, the pre-peroxisomal vesicles fuse with divided peroxisomes generated from preexisting mature peroxisomes. Peroxisome division requires Pex11 and a specific set of DRPs. In plants, retrograde trafficking from peroxisomes to the ER has been described (McCartney et al., 2005).")
33
A Pex19 alternatív szerepe a peroxiszómális
membránfehérjék (PMP-k) membránba illesztésében The role of Pex19 in peroxisome biogenesis and import of various PMPs has been clearly established in yeast and mammals, but its mechanism of action is still a matter of debate (Snyder et al., 1999; Sacksteder et al., 2000). Previous studies implicated Pex3 and Pex19 in the posttranslational insertion of PMPs. Pex19 serving as a chaperone binds and stabilizes newly synthesized mPTS-containing PMPs in the cytoplasm, and transports them to peroxisomes by docking to Pex3 present in the peroxisomal membrane (Muntau et al., 2003; Fang et al., 2004; Jones et al., 2004; Matsuzono and Fujiki, 2006; Matsuzono et al., 2006). However, subsequent studies in yeast show the requirement of Pex19 for the exit of most, if not all PMPs, including Pex3, from the ER (Fig. 2 B; Hoepfner et al., 2005; Lam et al., 2010; van der Zand et al., 2010; unpublished data). In the light of the Pex19-independent insertion of most PMPs into the ER and the role of Pex19 in mediating the budding of pre-peroxisomal vesicles, the role of Pex19 in the posttranslational import of PMPs is questionable for all PMPs that go to peroxisomes via the ER.
 membránba illesztésében. The role of Pex19 in peroxisome biogenesis and import of various PMPs has been clearly established in yeast and mammals, but its mechanism of action is still a matter of debate (Snyder et al., 1999; Sacksteder et al., 2000). Previous studies implicated Pex3 and Pex19 in the posttranslational insertion of PMPs. Pex19 serving as a chaperone binds and stabilizes newly synthesized mPTS-containing PMPs in the cytoplasm, and transports them to peroxisomes by docking to Pex3 present in the peroxisomal membrane (Muntau et al., 2003; Fang et al., 2004; Jones et al., 2004; Matsuzono and Fujiki, 2006; Matsuzono et al., 2006). However, subsequent studies in yeast show the requirement of Pex19 for the exit of most, if not all PMPs, including Pex3, from the ER (Fig. 2 B; Hoepfner et al., 2005; Lam et al., 2010; van der Zand et al., 2010; unpublished data). In the light of the Pex19-independent insertion of most PMPs into the ER and the role of Pex19 in mediating the budding of pre-peroxisomal vesicles, the role of Pex19 in the posttranslational import of PMPs is questionable for all PMPs that go to peroxisomes via the ER.")
34
A peroxiszóma komponensek továbbilehetséges
forrásai

35
Peroxiszómális fehérjék és a velük kapcsolatos emberi betegségek

36
Peroxiszómális betegségek
Mendeli öröklődés Zellweger szindróma 1 hónapon belüli halál Üres peroxiszómák Mutáns Pex5

37
Zellweger fibroblasztok Genetikailag módosított
Peroxiszóma nélküli Zellweger fibroblasztok Genetikailag módosított (Pex5 transzfektált) fibroblasztok peroxiszómákkal
 fibroblasztok. peroxiszómákkal.")
38
X-hez kötött újszülöttkori adrenoleukodystrophia (ALD)
16 hónapon belüli halál Abnormális oxidáció VLCFA felhalmozódás VLCFA = Very long chain fatty acid = nagyon hosszú láncú zsírsav ALDP = ALD protein = ABC transzporter (D1 típusú)
")
39
Zsírsav lebontás a peroxiszómában

40
22 évvel élt tovább az orvosok által jósoltnál!!
Lorenzo Odone ( ) 22 évvel élt tovább az orvosok által jósoltnál!!
 22 évvel élt tovább az orvosok által jósoltnál!!")
41
Lorenzo olaja mellett az ex vivo génterápia is egy távlati lehetőség

42
A ZELLWEGER SZINDRÓMA-SZERŰ RENDELLENESSÉGEKKEL KAPCSOLATOS PEX MUTÁCIÓK
GYAKORISÁG FUNKCIÓ % PEX Peroxisome biogenezis és PTS receptor reciklizálás a citoszolba PEX E3 ligáz; PTS receptor ubiquitináció PEX < PMP biogenezis és Pex19 receptor PEX A peroxiszómális mátrix protein import PTS1 receptora PEX Peroxiszóma biogenezis és PTS receptor reciklizálás a citoszolba PEX E3 ligáz; PTS receptor ubiquitináció PEX E3 ligáz; PTS receptor ubiquitináció PEX Peroxiszóálisl mátrix protein import PEX < A peroxiszómális mátrix protein import transzlokonjának egyik komponense PEX PMP biogenezis PEX < PMP biogenezis; pre-peroxiszómális vezikulumok lefűződése az ER- ról PEX A Pex6 peroxiszómális membrán receptora ZSS, a main subgroup of PBDs, is comprised of the following diseases: Zellweger syndrome, neonatal adrenoleukodystrophy, and infantile Refsum disease. Mutations in the PEX7 gene are responsible for the second PBD subgroup, called rhizomelic chondrodysplasia punctata type I.

43
Mitokondrium Peroxiszóma Endoszimbionta Endoszimbionta DNS és fehérje
szintézis (korlátozott) Nincs DNS és fehérje szintézis Részleges lipid és fehérje import Részleges lipid és teljes fehérje import Oxidációs folyamatok ATP termelés Nincs H2O2 termelés Oxidációs folyamatok Nincs ATP termelés H2O2 termelés A mtDNS anyai öröklődése Mendeli öröklődés
 Nincs DNS és fehérje. szintézis. Részleges lipid és. fehérje import. Részleges lipid és teljes. fehérje import. Oxidációs folyamatok. ATP termelés. Nincs H2O2 termelés. Oxidációs folyamatok. Nincs ATP termelés. H2O2 termelés. A mtDNS anyai. öröklődése. Mendeli öröklődés.")
44
Endoszimbionták = sejtorganellumok
Membrán nélkül - Ostor/Centriólum Membránnal Mitokondrium Peroxiszóma Kloroplasztisz

46
Az eukaryota ostor eredete I.
Cm= cell membrane= sejtmembrán; bw=bacterial cell wall= bakteriális sejtfal; bg= bacterial genome= bakteriális genom; bm= bacterial membrane = bakteriális sejtmembrán; evm= endosymbiotic membrane=endoszimbionta membrán; ea= extrusive apparatus7 extrúziós (kitaszító) apparátus; mt= microtubule= mikrotubulus
 apparátus; mt= microtubule= mikrotubulus.")
47
Az eukaryota ostor eredete II.

48
Az eukaryota ostor eredete III.
ad= axonemal dynein= axonémális (tengelyfonal) dinein
 dinein.")
49
Az ostor/csilló szerkezete

50
A centriólum és a bazális test szerkezete

51
Csillótípusok Mozgékony monocilium (9+0 tengelyfonal = axonéma) a Hensen- csomóban található, az ún. nodális áramlás létrehozásáért felelős. A jobb-bal testszimmetria létrehozásához nélkülözhetetlen. Mozgékony csillók (9+2 tengelyfonalas szerkezet) az epitélium feletti extracelluláris folyadék továbbításában vesznek részt a petevezetőben, az agy ependymájában, illetve a légutakban. A spermium ostora egy a mozgásra specializálódott mozgékony, megnyúlt (9+2) csilló. Nem-mozgékony csillók (9+0 tengelyfonal) a legtöbb nem-mozgó sejt felszínéről kinyúlva olyan környezeti szignálokat érzékelnek, mint pl. a folyadékáramlás és a folyadékösszetétel. A vese tubulusok epitéliuma és az epevezeték, valamint a hasnyálmirigy vezeték epitéliuma jó példák a monociliumok előfordulására. A chondrocyta és osteocyta monociliumok valószínüleg a feszülés/torzulás mértékét érzékelik. A fotoreceptor sejtek specializált nem-mozgékony (9+0) csillói, amelyek a kül- és beltagot kötik össze. A belső fül nem mozgékony 9+2 csillói (kinocilium; stereocilium). A monociliated cell is shown in the centre. Motile monocilia (9+0 axoneme, middle left panels) are found at the embryonic node and generate the nodal flow that is essential for determination of left–right body asymmetry. Multiple motile cilia (9+2 axonemal structure, top panels) that transport extracellular fluid along the epithelial surface are located on respiratory epithelial cells, brain ependymal cells and epithelial cells lining the fallopian tubes (panel reproduced with permission from Ref. 59 © (2005) Elsevier). The sperm flagellum (top right panel; co-stained with antibodies against the dynein heavy chain DNAH5, red) represents a specialized, elongated motile cilium (9+2) that confers motility. Non-motile monocilia (9+0, bottom panels) extend from the surface of most quiescent cells of the body and sense environmental signals such as fluid flow and/or fluid composition. Well-known examples are the monocilia of the tubular epithelia of the kidney, and the epithelia of the bile duct) and pancreatic ducts. The chondrocyte and osteocyte monocilia probably function to sense the amount of strain in bones. The connecting cilia of photoreceptor cells are specialized non-motile cilia (9+0) that connect the inner and outer segments. Non-motile 9+2 cilia (middle right panel) are found in the inner ear (kinocilium, red, arrowhead; stereocilia, green) (panel reproduced with permission from Ref. 10 © (2005) John Wiley & Sons, Inc.). Besides the four cilia types shown, there might be a high variability of the axonemal structures within vertebrates. In all panels, axonemes were stained (red or green) by indirect immunofluorescence using an antibody against the cilia-specific acetylated a-tubulin isoform. Nuclei were stained using Hoechst or 4',6-diamidino-2-phenylindole (DAPI).
 az epitélium feletti. extracelluláris folyadék továbbításában vesznek részt a petevezetőben, az agy ependymájában, illetve a légutakban. A spermium ostora egy a mozgásra specializálódott mozgékony, megnyúlt (9+2) csilló. Nem-mozgékony csillók (9+0 tengelyfonal) a legtöbb nem-mozgó sejt. felszínéről kinyúlva olyan környezeti szignálokat érzékelnek, mint pl. a. folyadékáramlás és a folyadékösszetétel. A vese tubulusok epitéliuma és az epevezeték, valamint a hasnyálmirigy vezeték epitéliuma jó példák a monociliumok előfordulására. A chondrocyta és osteocyta monociliumok valószínüleg a feszülés/torzulás mértékét érzékelik. A fotoreceptor sejtek specializált nem-mozgékony (9+0) csillói, amelyek a kül- és beltagot kötik össze. A belső fül nem mozgékony 9+2 csillói (kinocilium; stereocilium). A monociliated cell is shown in the centre. Motile monocilia (9+0 axoneme, middle left panels) are found at the embryonic node and generate the nodal flow that is essential for determination of left–right body asymmetry. Multiple motile cilia (9+2 axonemal structure, top panels) that transport extracellular fluid along the epithelial surface are located on respiratory epithelial cells, brain ependymal cells and epithelial cells lining the fallopian tubes (panel reproduced with permission from Ref. 59 © (2005) Elsevier). The sperm flagellum (top right panel; co-stained with antibodies against the dynein heavy chain DNAH5, red) represents a specialized, elongated motile cilium (9+2) that confers motility. Non-motile monocilia (9+0, bottom panels) extend from the surface of most quiescent cells of the body and sense environmental signals such as fluid flow and/or fluid composition. Well-known examples are the monocilia of the tubular epithelia of the kidney, and the epithelia of the bile duct) and pancreatic ducts. The chondrocyte and osteocyte monocilia probably function to sense the amount of strain in bones. The connecting cilia of photoreceptor cells are specialized non-motile cilia (9+0) that connect the inner and outer segments. Non-motile 9+2 cilia (middle right panel) are found in the inner ear (kinocilium, red, arrowhead; stereocilia, green) (panel reproduced with permission from Ref. 10 © (2005) John Wiley & Sons, Inc.). Besides the four cilia types shown, there might be a high variability of the axonemal structures within vertebrates. In all panels, axonemes were stained (red or green) by indirect immunofluorescence using an antibody against the cilia-specific acetylated a-tubulin isoform. Nuclei were stained using Hoechst or 4 ,6-diamidino-2-phenylindole (DAPI).")
52
Csilló típusok és a szervezeten belüli előfordulásuk
A monociliated cell is shown in the centre. Motile monocilia (9+0 axoneme, middle left panels) are found at the embryonic node and generate the nodal flow that is essential for determination of left–right body asymmetry. Multiple motile cilia (9+2 axonemal structure, top panels) that transport extracellular fluid along the epithelial surface are located on respiratory epithelial cells, brain ependymal cells and epithelial cells lining the fallopian tubes (panel reproduced with permission from Ref. 59 © (2005) Elsevier). The sperm flagellum (top right panel; co-stained with antibodies against the dynein heavy chain DNAH5, red) represents a specialized, elongated motile cilium (9+2) that confers motility. Non-motile monocilia (9+0, bottom panels) extend from the surface of most quiescent cells of the body and sense environmental signals such as fluid flow and/or fluid composition. Well-known examples are the monocilia of the tubular epithelia of the kidney, and the epithelia of the bile duct (panel reproduced with permission from Ref. 75 © (2006) American Physiological Society) and pancreatic ducts (panel reproduced with permission from Ref. 129 © (2006) Elsevier). The chondrocyte and osteocyte monocilia probably function to sense the amount of strain in bones. The connecting cilia of photoreceptor cells are specialized non-motile cilia (9+0) that connect the inner and outer segments. Non-motile 9+2 cilia (middle right panel) are found in the inner ear (kinocilium, red, arrowhead; stereocilia, green) (panel reproduced with permission from Ref. 10 © (2005) John Wiley & Sons, Inc.). Besides the four cilia types shown, there might be a high variability of the axonemal structures within vertebrates. In all panels, axonemes were stained (red or green) by indirect immunofluorescence using an antibody against the cilia-specific acetylated a-tubulin isoform. Nuclei were stained using Hoechst or 4',6-diamidino-2-phenylindole (DAPI).
 are found at the embryonic node and generate the nodal flow that is essential for determination of left–right body asymmetry. Multiple motile cilia (9+2 axonemal structure, top panels) that transport extracellular fluid along the epithelial surface are located on respiratory epithelial cells, brain ependymal cells and epithelial cells lining the fallopian tubes (panel reproduced with permission from Ref. 59 © (2005) Elsevier). The sperm flagellum (top right panel; co-stained with antibodies against the dynein heavy chain DNAH5, red) represents a specialized, elongated motile cilium (9+2) that confers motility. Non-motile monocilia (9+0, bottom panels) extend from the surface of most quiescent cells of the body and sense environmental signals such as fluid flow and/or fluid composition. Well-known examples are the monocilia of the tubular epithelia of the kidney, and the epithelia of the bile duct (panel reproduced with permission from Ref. 75 © (2006) American Physiological Society) and pancreatic ducts (panel reproduced with permission from Ref. 129 © (2006) Elsevier). The chondrocyte and osteocyte monocilia probably function to sense the amount of strain in bones. The connecting cilia of photoreceptor cells are specialized non-motile cilia (9+0) that connect the inner and outer segments. Non-motile 9+2 cilia (middle right panel) are found in the inner ear (kinocilium, red, arrowhead; stereocilia, green) (panel reproduced with permission from Ref. 10 © (2005) John Wiley & Sons, Inc.). Besides the four cilia types shown, there might be a high variability of the axonemal structures within vertebrates. In all panels, axonemes were stained (red or green) by indirect immunofluorescence using an antibody against the cilia-specific acetylated a-tubulin isoform. Nuclei were stained using Hoechst or 4 ,6-diamidino-2-phenylindole (DAPI).")
53
Testszimmetria és a monociliumok
a | Schematic illustration of normal left–right body asymmetry (situs solitus) and five laterality defects that affect the lungs, heart, liver, stomach and spleen. By their vigorous circular movements, motile monocilia at the embryonic node generate a leftward flow of extra-embryonic fluid (nodal flow). b | The nodal vesicular parcel (NVP) model predicts that vesicles filled with morphogens (such as sonic hedgehog and retinoic acid) are secreted from the right side of the embryonic node and transported to the left side by nodal flow, where they are smashed open by force. The released contents probably bind to specific transmembrane receptors in the axonemal membrane of cilia on the left side. The consequent initiation of left-sided intracellular Ca2+ release induces downstream signalling events that break bilaterality. In this model, the flow of extra-embryonic fluid is not detected by cilia-based mechanosensation. c | In the two-cilia model, non-sensing motile cilia in the centre of the node create a leftward nodal flow that is mechanically sensed through passive bending of non-motile sensory cilia at the periphery of the node. Bending of the cilia on the left side leads to a left-sided release of Ca2+ that initiates the establishment of body asymmetry.
 and five laterality defects that affect the lungs, heart, liver, stomach and spleen. By their vigorous circular movements, motile monocilia at the embryonic node generate a leftward flow of extra-embryonic fluid (nodal flow). b | The nodal vesicular parcel (NVP) model predicts that vesicles filled with morphogens (such as sonic hedgehog and retinoic acid) are secreted from the right side of the embryonic node and transported to the left side by nodal flow, where they are smashed open by force. The released contents probably bind to specific transmembrane receptors in the axonemal membrane of cilia on the left side. The consequent initiation of left-sided intracellular Ca2+ release induces downstream signalling events that break bilaterality. In this model, the flow of extra-embryonic fluid is not detected by cilia-based mechanosensation. c | In the two-cilia model, non-sensing motile cilia in the centre of the node create a leftward nodal flow that is mechanically sensed through passive bending of non-motile sensory cilia at the periphery of the node. Bending of the cilia on the left side leads to a left-sided release of Ca2+ that initiates the establishment of body asymmetry.")
54
A spermiohisztogenezis lépései

55
dynein karok és szabálytalan elrendeződésű mikrotubulus párok.
TEM: egy normális csilló kültagjának keresztmetszete a 9 x mikrotubulussal. A külső és a belső dynein karok jól láthatók (nyilak). Egy primer ciliáris dyskinesiában szenvedő beteg csillójának keresztmetszete. Hiányzó dynein karok és szabálytalan elrendeződésű mikrotubulus párok.
. Egy primer ciliáris dyskinesiában szenvedő beteg csillójának keresztmetszete. Hiányzó. dynein karok és szabálytalan elrendeződésű mikrotubulus párok.")
56
Csillókkal kapcsolatos betegségek
Emberi betegség Gén (protein) Öröklődés Sejtes menet lokalizáció Primer ciliáris diszkinezia (PCD) DNAH1, DNAH5 Autoszómális recesszív Mozgékony csilló bazális test Policisztás vesebetegség (PKD) Polycystin1, polycystin2, fibrocystin Autoszómális recesszív Primer csilló/ bazális test Nephronophthisis (NPHP) Nephrocystin, inversin Autoszómális recesszív Csilló/bazális test Bardet–Biedl szindróma (BBS) BBS1, BBS4, BBS7, BBS8, BBS10 Autoszómális recesszív Csilló/bazális test Retinits pigmentosa (RP) RP1, RPGR Autoszómális recesszív/ Fotoreceptor és csilló X-hez kötött recesszív junkció Vastagbélrák Gli3 ……………………. Csilló/bazális test Meckel–Gruber szindróma MKS1, MKS3 Autoszómális recesszív Bazális test Száj-arc-kéz szindróma ODF1 X-hez kötött recesszív Bazális test Csillókkal kapcsolatos betegségek Abbreviations: BBS, Bardet-Biedl syndrome; NPHP, nephronophthisis; PCD, primary ciliary dyskinesia; PKD, polycystic kidney disease; RP, retinits pegmentosa.
 Öröklődés- Sejtes menet lokalizáció. Primer ciliáris diszkinezia (PCD) DNAH1, DNAH5 Autoszómális recesszív Mozgékony csilló bazális test. Policisztás vesebetegség (PKD) Polycystin1, polycystin2, fibrocystin Autoszómális recesszív Primer csilló/ bazális test. Nephronophthisis (NPHP) Nephrocystin, inversin Autoszómális recesszív Csilló/bazális test. Bardet–Biedl szindróma (BBS) BBS1, BBS4, BBS7, BBS8, BBS10 Autoszómális recesszív Csilló/bazális test. Retinits pigmentosa (RP) RP1, RPGR Autoszómális recesszív/ Fotoreceptor és csilló X-hez kötött recesszív junkció. Vastagbélrák Gli3 ……………………. Csilló/bazális test. Meckel–Gruber szindróma MKS1, MKS3 Autoszómális recesszív Bazális test. Száj-arc-kéz szindróma ODF1 X-hez kötött recesszív Bazális test. Csillókkal kapcsolatos betegségek. Abbreviations: BBS, Bardet-Biedl syndrome; NPHP, nephronophthisis; PCD, primary ciliary dyskinesia; PKD, polycystic kidney disease; RP, retinits pegmentosa.")
57
Csillóbetegségekben érintett fehérjék és kapcsolatuk a csilló-centroszóma-komplex-szel (CCC)
")
59
Az összekötő csillók alapvető fontosságúak a
fotoreceptorok élettanában The connecting 9+0 cilia of photoreceptors represent specialized cilia that are the sole transport corridor between the outer and inner photoreceptor segments. These cilia are essential in photoreceptor physiology and, therefore, their dysfunction contributes to retinal degeneration. a | Schematic illustration of a photoreceptor cell and its substructures. b | Immunofluorescence staining of retinal sections using specific markers for ciliary subcompartments. Antibodies against acetylated -tubulin mark the axoneme (green, left panel). The transition zone was visualized using antibodies against nephrocystin (red, left and right panels), and the basal bodies were stained using antibodies against the pericentriolar marker -tubulin (green, right panel). c | Electron microscopy image of a retina showing the localization of the photoreceptor-connecting cilium between the outer and inner segments. d | Schematic illustration of the physiological function of a connecting cilium. Biosynthesis products from the inner segment and turnover products from the outer segment are shuttled through the connecting cilium by the IFT machinery. The localization of several proteins implicated in retinal diseases is indicated: RPGR and nephrocystin are found in the transition zone, usherin is located in the ciliary membrane and BBS proteins are found in basal bodies. BBS, Bardet–Biedl syndrome; BM, Bruch's membrane; IFT, intraflagellar transport; INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; RPE, retinal pigment epithelium; RPGR, retinitis pigmentosa guanosine triphosphatase (GTPase) regulator. Part a modified with permission from Ref. 100 © (2006) Elsevier.
. The transition zone was visualized using antibodies against nephrocystin (red, left and right panels), and the basal bodies were stained using antibodies against the pericentriolar marker -tubulin (green, right panel). c | Electron microscopy image of a retina showing the localization of the photoreceptor-connecting cilium between the outer and inner segments. d | Schematic illustration of the physiological function of a connecting cilium. Biosynthesis products from the inner segment and turnover products from the outer segment are shuttled through the connecting cilium by the IFT machinery. The localization of several proteins implicated in retinal diseases is indicated: RPGR and nephrocystin are found in the transition zone, usherin is located in the ciliary membrane and BBS proteins are found in basal bodies. BBS, Bardet–Biedl syndrome; BM, Bruch s membrane; IFT, intraflagellar transport; INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; RPE, retinal pigment epithelium; RPGR, retinitis pigmentosa guanosine triphosphatase (GTPase) regulator. Part a modified with permission from Ref. 100 © (2006) Elsevier.")
60
Endoszimbionták = sejtorganellumok
Membrán nélkül - Ostor/Centriólum Membránnal Mitokondrium Peroxiszóma Kloroplasztisz

63
(korábbi nevén kékalga) elektronmikroszkópos képe
A zöld színtest és a cianobaktérium (korábbi nevén kékalga) elektronmikroszkópos képe
 elektronmikroszkópos képe.")
64
A mitokondrium és a színtest összehasonlítása
2 membránrendszer! Külső Belső 3 membránrendszer! Külső Belső Tilakoid

65
Szerkezet

66
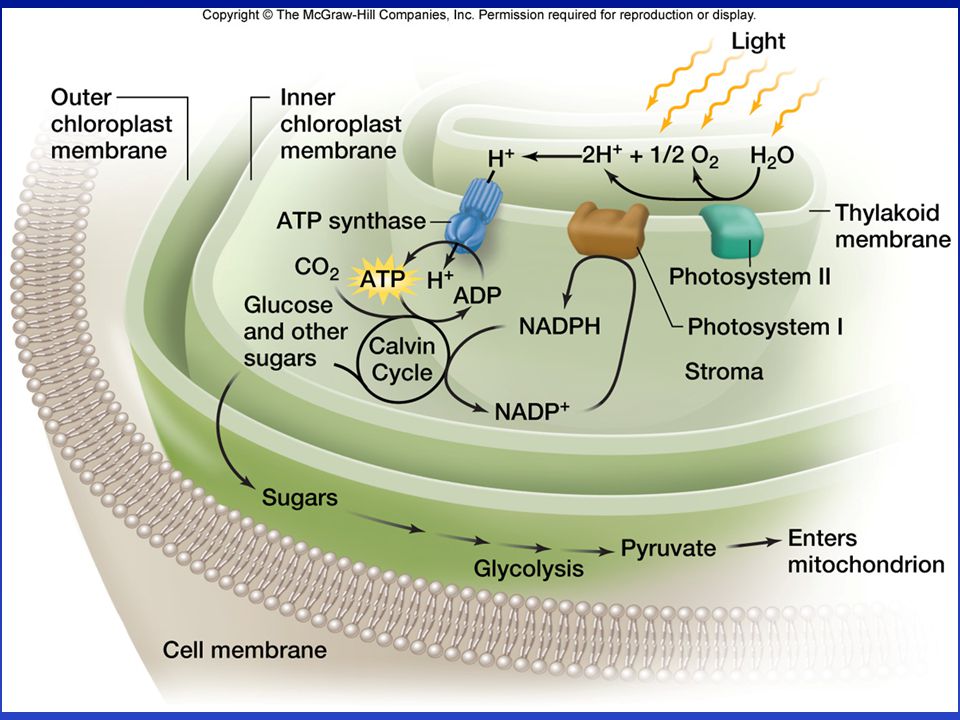
Funkció: Fotoszintézis 12H2O + 6CO2 + fény = (CH2O)6 + 6O2
De a legtöbb citoszol ATP a növényekben is a mitokondriumból származik. Típusai lamelláris (algák) granumos (szárazföldi növények)
 granumos. (szárazföldi növények)")
67
A thylakoid membrán funkciója
Fotoszintetikus elektrontranszfer, ATP és NADPH2 szintézis („fényreakció”) H2O +NADP+ + 2Pi + 2ADP + fény = 1/2O2 + NADPH2 +2ATP + 2H2O
 H2O +NADP+ + 2Pi + 2ADP + fény = 1/2O2 + NADPH2 +2ATP + 2H2O.")
69
A sztróma funkciója Purin, pirimidin és részben aminosav szintézis Replikáció, transzkripció és transzláció Szén-dioxid megkötés (ribulóz bifoszfát karboxiláz) és cukor szintézis („sötét reakció”) a Calvin-Benson ciklusban: 6CO2 +18ATP + 12NADPH + 12H+ = (CH2O)6 + 18ADP + 18Pi +12NADP+
 és cukor szintézis („sötét reakció ) a Calvin-Benson ciklusban: 6CO2 +18ATP + 12NADPH + 12H+ = (CH2O)6 + 18ADP + 18Pi +12NADP+")
70
A színtestek képződése
Osztódás Ketté hasadás Aszinkron a magi osztódással, de szabályozott! A DNS-ek random replikációja Lipidek eredete: színtest Protein import Hasonló a mitokondriálishoz

71
A genom (DNS) elhelyezkedése a mitokondriumban
és a zöld színtestben

72
Az Anopheles gambiae a leggyakoribb
maláriát terjesztő szúnyog(moszkitó)
")
73
A malária parazita életciklusa

74
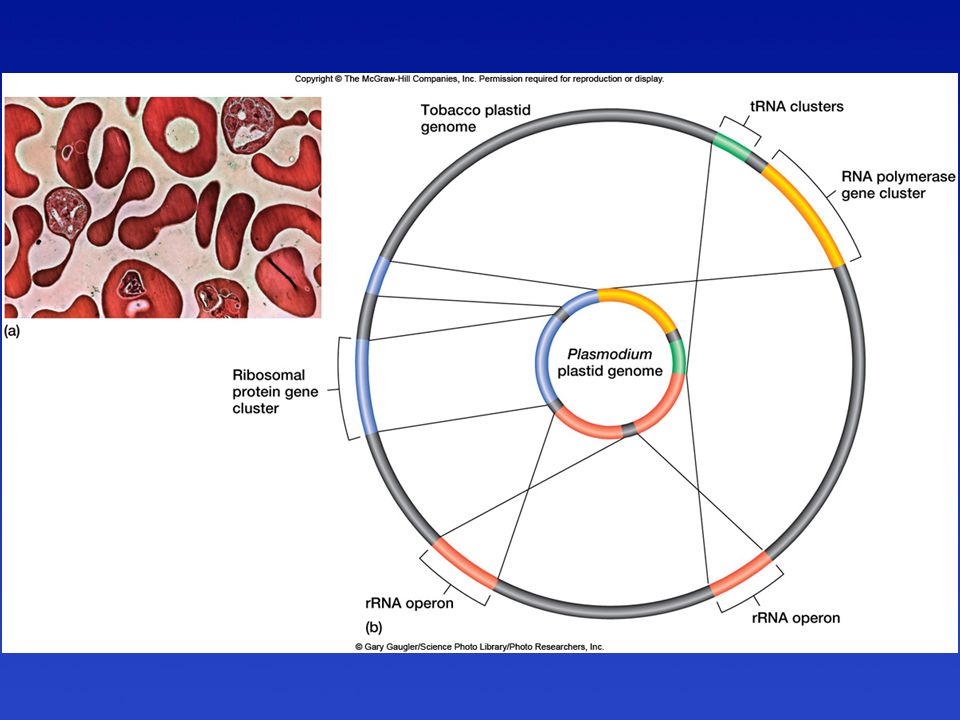
Plasmodium falciparum parazitával fertőzött maláriás emberi vér
A nyilak a parazita különböző fejlődési stádiumaira mutatnak

76
Kloroplasztisz genomok
összehasonlítása

78
Miért van csökevényes színtest (plasztid) a Plasmodium parazitában?
Feltételezések szerint egy parazita életmódot folytató tengeri Dinoflagellátától, egy egysejtű páncélos ostorostól származik, amelynek ostora, mitokondriuma és színtestje is volt A parazita életmód következtében ezek elcsökevényesedtek illetve eltűntek

79
A plasztid DNS terápiás célpont lehet a malária elleni védekezésben

Hasonló előadás