Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
? Kialakulás - A peroxiszóma és a mitokondrium bekebelezése előtt
Az eukarióta sejtváz/ citoszkeleton Filamentumokból, mikrotubulusokból valamint ezek működésében résztvevő és szabályozó molekulák összessége Kialakulás - A peroxiszóma és a mitokondrium bekebelezése előtt ? - méret növekedést követő stabilitás, kompartmentalizáció De elemei : már a prokariótákban

2
Az eukarióta sejtváz komponensei
Mikrofilamentumok alapfehérje: aktin Mikrotubulusok Alapfehérje: tubulin Intermedier filamentumok Sejt/szövet specifikus Kivétel: laminok

3
Mikrotubulusok mikrotubulusok: zöld aktin: piros
migration.wordpress.com/.../

4
A prokariótákban: tubulin analógok
www2.mrc-lmb.cam.ac.uk/SS/Lowe_J/ Tubulin analóg fehérjék D szerkezet hasonló

5
Baktériumok kettéfűződésében szerep: Z ring (Z gyűrű)
Tubulin analóg FtsZ Baktériumok kettéfűződésében szerep: Z ring (Z gyűrű) In step 1, newborn cells grown at low growth rates contain a single, non-replicating chromosome in a structure known as a nucleoid. Soon after chromosome replication initiates, the replication origins (oriC) move towards the cell poles until both daughter chromosomes are segregated (step 2). Near the end of this process, the FtsZ protein assembles into the Z ring on the inner face of the cytoplasmic membrane (light green ring) at the cell centre, marking the future division site (step 3). In step 4, the Z ring recruits at least ten membrane-associated proteins (for details, see Fig. 6) to assemble the cell-division protein machinery (dark green ring). This machinery synthesizes the division septum, which consists of cell-wall material, with the Z ring at the leading edge of membrane invagination. Contraction of the Z ring and constriction of the outer membrane follow (step 5). The result is the production of two separate newborn daughter cells (step 6).
 In step 1, newborn cells grown at low growth rates contain a single, non-replicating chromosome in a structure known as a nucleoid. Soon after chromosome replication initiates, the replication origins (oriC) move towards the cell poles until both daughter chromosomes are segregated (step 2). Near the end of this process, the FtsZ protein assembles into the Z ring on the inner face of the cytoplasmic membrane (light green ring) at the cell centre, marking the future division site (step 3). In step 4, the Z ring recruits at least ten membrane-associated proteins (for details, see Fig. 6) to assemble the cell-division protein machinery (dark green ring). This machinery synthesizes the division septum, which consists of cell-wall material, with the Z ring at the leading edge of membrane invagination. Contraction of the Z ring and constriction of the outer membrane follow (step 5). The result is the production of two separate newborn daughter cells (step 6).")
6
Prokarióta eredet igazolása
Színtest és mitokondrium: megtalálhatók a bakteriális rendszer egyes komponensei Different types of cells or organelles and their use of FtsZ or alternative proteins for fission are shown. In euryarchaea and many bacteria, FtsZ (green) localizes to the inner side of the inner membrane as the Z ring. Crenarchaea and some bacteria lack FtsZ, so some other protein must localize as a ring at the site of division (yellow). In plastids and mitochondria, dynamin or dynamin-like protein rings (red) localize to the cytoplasmic face. These organelles also contain a dividing ring (blue) and, in the case of chloroplasts and some primitive mitochondria, an innermost ring of FtsZ (green). Most mitochondria, including those of fungi and animals, lack FtsZ and a detectable dividing ring, but still rely on dynamin on the cytosolic face for fission (red).
 localizes to the inner side of the inner membrane as the Z ring. Crenarchaea and some bacteria lack FtsZ, so some other protein must localize as a ring at the site of division (yellow). In plastids and mitochondria, dynamin or dynamin-like protein rings (red) localize to the cytoplasmic face. These organelles also contain a dividing ring (blue) and, in the case of chloroplasts and some primitive mitochondria, an innermost ring of FtsZ (green). Most mitochondria, including those of fungi and animals, lack FtsZ and a detectable dividing ring, but still rely on dynamin on the cytosolic face for fission (red).")
7
Mikrotubulusok elrendeződése:
Mikrotubulusok típusai: Citoplazmatikus mikrotubulusok: Egyediek, kapcsolódás más citoszkeletális komponensekkel Szerveződés: sejtközpont (centriólum, MTOC) Csilló/ostor/bazális test/centriólum mikrotubulusok: Csoportokban (párosával – doublet, hármasával – triplet) „Állandósult” sejtfelszíni és sejtbeni képletek Mikrotubulusok elrendeződése: - függ a sejt típusától: polarizált sejt stb…. - függ a sejtciklus szakaszától: profázisos sejt stb….
 Csilló/ostor/bazális test/centriólum mikrotubulusok: Csoportokban (párosával – doublet, hármasával – triplet) „Állandósult sejtfelszíni és sejtbeni képletek. Mikrotubulusok elrendeződése: - függ a sejt típusától: polarizált sejt stb…. - függ a sejtciklus szakaszától: profázisos sejt stb….")
8
MTOC:mikrotubule organizer center
Mikrotubulusok elrendeződése az eukarióta sejtekben Interphase Dividing cell Neuron centrosome Basal body Cilium Mitotic spindle axon MTOC:mikrotubule organizer center többféle elrendezésben szervezi a mikrotubulusokat -interfázisos (nem osztódó sejtben) -neuronokban : speciális az axonban -osztódó sejtben mitótikus orsó -csilló ill. ostor
 -neuronokban : speciális az axonban. -osztódó sejtben mitótikus orsó. -csilló ill. ostor.")
9
Citoplazmatikus mikrotubulusok

10
Mitótikus orsó mikrotubulusai

11
csilló Basal body google.com/.../BDSUCIwoQtPHboPYi

12
polaritás (+ és – vég) polimerizáció tubulin heterodimerek ( és )
Mikrotubulusok felépítése polimerizáció tubulin heterodimerek ( és ) GTP és GDP kötő képesség polaritás (+ és – vég)
 GTP és GDP kötő képesség. polaritás (+ és – vég)")
13
Mikrotubulusok jellemzői: Dinamikus instabilitás
GTP sapka szerepe: növekvő mikrotubulus + végén GTP-t kötő egységek belül GDP-t kötő egységek raven.zoology.washington.edu/.../index.html GTP sapka elvesztése mikrotubulus depolimerizáció

14
Mikrotubulusok jellemzői:„Taposómalom” treadmill jelensége
+ vég, növekvő vég polimerizáció - vég, a fogyó vég depolimerizáció Stabilitás biztosítása :MAPs (microtubule associated proteins)
")
15
merőleges elrendezésben Pericentrioláris mátrixban:
Citoplazmatikus mikrotubulusok szervezése: MTOC (centriólum, sejtközpont) - mikrotubulusokból épül fel g-tubulin 9x3 microtubulus A,B,C 2 centriolum egymásra merőleges elrendezésben Pericentrioláris mátrixban: -tubulin mikrotubulusok kiindulási (nukleációs helye) (Brinkley, B.R. Encyclop. Neurosci. 1987, 665)
 - mikrotubulusokból épül fel. g-tubulin. 9x3 microtubulus A,B,C. 2 centriolum egymásra. merőleges elrendezésben. Pericentrioláris mátrixban: -tubulin mikrotubulusok kiindulási (nukleációs helye) (Brinkley, B.R. Encyclop. Neurosci. 1987, 665)")
16
MTOC :Mikrotubulusok nukleációs helye
γ-Tubulin Ring Complex: 3D Tomographic reconstructions negative stain (γTuSC outlined) a pericentrioláris mátrixban -tubulin gyűrű nukleációs hely vég felöl polimerizáció Stabilizáció : pl. sapkaképző fehérjékkel (MAPs –ok egyik képviselője)
 a pericentrioláris mátrixban -tubulin gyűrű nukleációs hely. vég felöl polimerizáció. Stabilizáció : pl. sapkaképző fehérjékkel (MAPs –ok egyik képviselője)")
17
MAPs fehérjék (mikrotubulus associated proteins)
MT polimerizáció/depolimerizáció: stathmin: tubulin egységek leválása, depolimerizáció serkentése CRMP2 (collapsin response mediator protein): polimerizáció serkentése, dimer egységek beépülése a + végen 2. mikrotubulusok stabilizációja: tau mikrotubulusok dinamikus állapota: MAP1b mikrotubulusok kötegelése: MAP1c APC (adenomateous poliposis coli) protein: MT + véghez kötődő fehérjékkel komplexet képez és stabilizálja a növekedést 3. Motor fehérjék: mechano-kémiai enzimek 2 család: kinezin és dinein
: polimerizáció serkentése, dimer egységek beépülése a + végen. 2. mikrotubulusok stabilizációja: tau. mikrotubulusok dinamikus állapota: MAP1b. mikrotubulusok kötegelése: MAP1c. APC (adenomateous poliposis coli) protein: MT + véghez kötődő fehérjékkel komplexet képez és stabilizálja a növekedést. 3. Motor fehérjék: mechano-kémiai enzimek. 2 család: kinezin és dinein.")
18
mikrotubulus asszociált fehérjék
MAPs Szerkezeti fehérjék Tau, MAP1, MAP2 Motor fehérjék kinezin, dinein család =learnmore.genetics Szerep: sejtalkotók, vezikulumok mozgatása Szerep: mikrotubulusok stabilizálása

19
Figure 3: The Sf9 cell bodies have a round shape, with diameter of 20 µm. After expression of tau protein they develop a single process per cell, with uniform diameter of 1-2 µm and up to 100 µm long. from: Biernat and Mandelkow, Molec.Biol.Cell., 10, ,1999.

20
Citoplazmatikus dinein szerkezete
…szerkezeti hasonlóság más motor fehérjéhez Nehéz lánc: globuláris fej + farok Könnyű lánc: LC8: reguláció Intermedier lánc: IC

21
Kinezin és dinein működése: séta a mikrotubulusokon
cargo Globuláris fej: ATP kötés és bontás –séta a mikrotubuluson rövidebb (dinein) és hosszabb (kinezin) farok: cargo megkötése Kinesin: + vég motor Dinein: - vég motor
 és hosszabb (kinezin) farok: cargo megkötése. Kinesin: + vég motor. Dinein: - vég motor.")
22
Nature Reviews Neuroscience 6, 201-214 (March 2005)
Neuronok axonjaiban vezikuláris transzport Nature Reviews Neuroscience 6, (March 2005) Molecular motors and mechanisms of directional transport in neurons Nobutaka Hirokawa & Reiko Takemura
 Molecular motors and mechanisms of directional transport in neurons. Nobutaka Hirokawa & Reiko Takemura.")
23
Mitótikus orsó mikrotubulusai:osztódó sejtek
Centriólumok duplikációját követően a két pólus között mikrotubulusok speciális elrendeződése: mitótikus orsó

24
Mitótikus orsó szerkezete
3 féle MT: asztrális, kinetokor és poláris mikrotubulusok kinetokor: kromoszómák kromatidáinak mozgatása anafázisban (A) asztrális és poláris: a sejt elongációját segíti anafázisban (B)
 asztrális és poláris: a sejt elongációját segíti anafázisban (B)")
25
Microtubulus polimerizáció gátlás Mikrotubulus stabilizáció
Tiszafa Őszi kikerics Colchicin Taxol Microtubulus polimerizáció gátlás Mikrotubulus stabilizáció

26
Mikrotubulus „mérgek”
A few molecules of vinblastine bound to high-affinity sites at the microtubule plus end suffice to suppress microtubule dynamics. b | Colchicine forms complexes with tubulin dimers and copolymerizes into the microtubule lattice, suppressing microtubule dynamics. c | A microtubule cut away to show the interior surface is shown. Paclitaxel binds along the interior surface of the microtubule, suppressing its dynamics.

27
Próba: anti tumor szer: de rengeteg mellékhatás
COLCHICINE TAB MYM Reg: No. R 0808 A 3305 Composition: Colchicine 0.6 mg Description: White colour tab, blister pack of 10 x 10's Indications: Symptomatic relief of acute attacks of acute gouty arthritis and for prophylaxis against repeat attacks. Dosage & Administration: Initial 2 tab followed by 1 tab, every 1-2 hours until pain relief or nausea, vomiting or diarrhoea occur. utp.htm Köszvény elleni szer (granulocita mozgást gátolja) és íngyulladást csökkentő hatás Próba: anti tumor szer: de rengeteg mellékhatás commons.wikimedia.org/wiki/Image:Colchicine.svg
 és íngyulladást csökkentő hatás. Próba: anti tumor szer: de rengeteg mellékhatás. commons.wikimedia.org/wiki/Image:Colchicine.svg.")
28
Paclitaxel : mitózis inhibitor és tumor ellenes terápiában használatos
Hatóanyag: taxol Paclitaxel : mitózis inhibitor és tumor ellenes terápiában használatos tüdő, petefészek és emlőrákban, fej-nyakrákban és Kaposi szarkómában www-jmg.ch.cam.ac.uk/.../misc/taxol.html

29
* Stabilizált mikrotubulusokból felépülő képletek: csilló és ostor
„klasszikus csilló” (sok van) : mozgó: pl. trahea, agy, petevezeték, belső fül nem mozgó pl. belső fül stereocilium alapszerkezet: 9+2 (9x2+2) * Ostor: mozgó pl. spermium: speciális meghosszabbodott mozgó csilló/csillók Monocilium: egy csilló van, amely lehet: mozgó pl. nodális cilium - embrió * nem mozgó pl. vese szem –retina fotoreceptor alapszerkezet: (9x2+0) Töbféle csilló van: - mozgó (kinocilium), nem mozgó pl. stereocilium
 : mozgó: pl. trahea, agy, petevezeték, belső fül. nem mozgó pl. belső fül stereocilium. alapszerkezet: 9+2 (9x2+2) * Ostor: mozgó pl. spermium: speciális meghosszabbodott mozgó csilló/csillók. Monocilium: egy csilló van, amely lehet: mozgó pl. nodális cilium - embrió * nem mozgó pl. vese. szem –retina fotoreceptor. alapszerkezet: (9x2+0) Töbféle csilló van: - mozgó (kinocilium), nem mozgó pl. stereocilium.")
30
Trahea: többmagsoros, csillós hengerhám
SEM: csilló Trahea: többmagsoros, csillós hengerhám TEM: axonéma szerkezet

31
Bazális test A csilló ill. ostor MTOC tubulin: a csilló és ostor
Szerkezete: megegyezik a centriólum szerkezetével tubulin: a csilló és ostor mikrotubulusainak nukleációs helye

32
Csilló szerkezet: (klasszikus) 9 pár +2 mikrotubulus
A csilló un. axonéma szerkezete perifériás mikrotubulus párok (9) és egy centrális pár +2 9+0 szerkezet: hiányzik a centrális mikrotubulus pár átmeneti zóna és monociliumok bazális test szerkezete: mint MTOC, ugyanaz mint a centriólum szerkezete 9x3
 és egy centrális pár szerkezet: hiányzik a centrális mikrotubulus pár. átmeneti zóna és monociliumok. bazális test szerkezete: mint MTOC, ugyanaz mint a. centriólum szerkezete 9x3.")
33
szerkezeti: küllők ~ 17 polipeptid
Csilló: tubulin + MAPs szerkezeti: küllők ~ 17 polipeptid central sheat (centrális hüvely) fehérjéi Mikrotubulus párokat összekötő: nexin Motor protein: Csilló dinein Külső és belső dinein karok Nehéz lánc (HC) Intermedier lánc (IC) Könnyű lánc (LC)
 fehérjéi. Mikrotubulus párokat összekötő: nexin. Motor protein: Csilló dinein. Külső és belső. dinein karok. Nehéz lánc (HC) Intermedier lánc (IC) Könnyű lánc (LC)")
34
Hogyan mozog a csilló?

35
Csilló lecsapás – aktív (ATP-t igénylő), a felemelkedés passzív
, a felemelkedés passzív")
36
Csillós mozgás mechanizmus: mikrotubulus sliding
Sliding (elcsúszás) Bending (lecsapás)
 Bending (lecsapás)")
37
Mikrotubulus „sliding” (elcsúszás)
")
38
Ostoros mozgás –szakaszosan ismétlődő
csillós mozgás

39
Speciális csillók és funkcióik
Taupátia és ciliopátiák Mikrotubulus asszociált fehérjékkel kapcsolatos: taupáthiák: MAPT mutációk - taupáthia mint egy komplex betegség része: Alzheimer kór Mikrotubulusból felépülő struktúra: csilló betegségei - ciliopáthiák

40
hedgehogs ? Sonic hedgehog

41
Mit csinálhat egy csilló ha nem mozog?
Hedgehog (Hh) signalling in cilia involves the intraflagellar transport machinery, which moves components of the Hh signalling pathway to their functional sites. The transcription factors glioma (GLI) and suppressor of fused (SUFU) are transported to the ciliary tip. GLI is processed to create a transcriptional repressor, which is transported back to the cell body (left panel). On Hh ligand binding to its receptor patched-1 (PTCH1), smoothened (SMO) is released and transported to the ciliary tip, where it turns off GLI processing by interacting with SUFU. The activator form of the GLI transcription factor is transported to the cell body and enters the nucleus where it induces the expression of genes, such as those involved in renal patterning (PAX2, SALL1), cell-cycle regulation (CCND1 (cyclin D1), N-MYC) and GLI family members themselves (GLI1, GLI2) and PTCH1 (self-induction). Embrionális fejlődés egyik szignalizációs útvonala : Hedgehog (Hh útvonal) A monocilium szerepe: intraflagelláris transzporttal szállítja a szignál útvonal komponenseit a helyükre (GLI, SUFU) – szignál nélkül a GLIR formája keletkezik és nincs transzkripció (represszor funkció) Szignál: Hh bekötődik a receptorához (PTCH1), ennek hatására az SMO fehérje leválik, felszállítódik a csillóban, a SUFU-val reagál és ennek hatására GLIA a GLI aktivátor formája keletkezik. Ez intraflagelláris transzporttal visszaszállítódik a sejtbe, bejut a sejtmagba és specifikus transzkripciós faktorként génexpressziót indukál.
 signalling in cilia involves the intraflagellar transport machinery, which moves components of the Hh signalling pathway to their functional sites. The transcription factors glioma (GLI) and suppressor of fused (SUFU) are transported to the ciliary tip. GLI is processed to create a transcriptional repressor, which is transported back to the cell body (left panel). On Hh ligand binding to its receptor patched-1 (PTCH1), smoothened (SMO) is released and transported to the ciliary tip, where it turns off GLI processing by interacting with SUFU. The activator form of the GLI transcription factor is transported to the cell body and enters the nucleus where it induces the expression of genes, such as those involved in renal patterning (PAX2, SALL1), cell-cycle regulation (CCND1 (cyclin D1), N-MYC) and GLI family members themselves (GLI1, GLI2) and PTCH1 (self-induction). Embrionális fejlődés egyik szignalizációs útvonala : Hedgehog (Hh útvonal) A monocilium szerepe: intraflagelláris transzporttal szállítja a szignál útvonal komponenseit a helyükre (GLI, SUFU) – szignál nélkül a GLIR formája keletkezik és nincs transzkripció (represszor funkció) Szignál: Hh bekötődik a receptorához (PTCH1), ennek hatására az SMO fehérje leválik, felszállítódik a csillóban, a SUFU-val reagál és ennek hatására GLIA a GLI aktivátor formája keletkezik. Ez intraflagelláris transzporttal visszaszállítódik a sejtbe, bejut a sejtmagba és specifikus transzkripciós faktorként génexpressziót indukál.")
42
Fotoreceptor sejtek: kültag és beltag
The connecting 9+0 cilia of photoreceptors represent specialized cilia that are the sole transport corridor between the outer and inner photoreceptor segments. These cilia are essential in photoreceptor physiology and, therefore, their dysfunction contributes to retinal degeneration. a | Schematic illustration of a photoreceptor cell and its substructures. b | Immunofluorescence staining of retinal sections using specific markers for ciliary subcompartments. Antibodies against acetylated -tubulin mark the axoneme (green, left panel). The transition zone was visualized using antibodies against nephrocystin (red, left and right panels), and the basal bodies were stained using antibodies against the pericentriolar marker -tubulin (green, right panel). c | Electron microscopy image of a retina showing the localization of the photoreceptor-connecting cilium between the outer and inner segments. d | Schematic illustration of the physiological function of a connecting cilium. Biosynthesis products from the inner segment and turnover products from the outer segment are shuttled through the connecting cilium by the IFT machinery. The localization of several proteins implicated in retinal diseases is indicated: RPGR and nephrocystin are found in the transition zone, usherin is located in the ciliary membrane and BBS proteins are found in basal bodies. BBS, Bardet–Biedl syndrome; BM, Bruch's membrane; IFT, intraflagellar transport; INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; RPE, retinal pigment epithelium; RPGR, retinitis pigmentosa guanosine triphosphatase (GTPase) regulator. Part a modified with permission from Ref. 100 © (2006) Elsevier. Fotoreceptor sejtek: kültag és beltag Kettő között a kapcsolat: összekötő csilló szegment amely az intraflagelláris transzportot végzi a két rész között
. The transition zone was visualized using antibodies against nephrocystin (red, left and right panels), and the basal bodies were stained using antibodies against the pericentriolar marker -tubulin (green, right panel). c | Electron microscopy image of a retina showing the localization of the photoreceptor-connecting cilium between the outer and inner segments. d | Schematic illustration of the physiological function of a connecting cilium. Biosynthesis products from the inner segment and turnover products from the outer segment are shuttled through the connecting cilium by the IFT machinery. The localization of several proteins implicated in retinal diseases is indicated: RPGR and nephrocystin are found in the transition zone, usherin is located in the ciliary membrane and BBS proteins are found in basal bodies. BBS, Bardet–Biedl syndrome; BM, Bruch s membrane; IFT, intraflagellar transport; INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; RPE, retinal pigment epithelium; RPGR, retinitis pigmentosa guanosine triphosphatase (GTPase) regulator. Part a modified with permission from Ref. 100 © (2006) Elsevier. Fotoreceptor sejtek: kültag és beltag. Kettő között a kapcsolat: összekötő csilló szegment. amely az intraflagelláris transzportot végzi a két rész között.")
43
Table I. Human ciliary disease genes and their cell biological functions
Cellular function Protein localization PKD PKD1 PKHD1 Mechanosensing Unknown Cilia Basal body Primary ciliary dyskinesis Bronchiectasis Chronic sinusitis Situs inversus DNAH5 DNAI1 Ciliary motility Outer dynein arms Bardet-Biedl syndrome Obesity Retinal degeneration Cystic kidneys BBS1-12 Ciliogenesis Basal body and IFT complexes Meckel-Gruber syndrome Brain malformation Polydactyly Cep290 MKS1 MKS3 Ciliary membrane Oral-Facial-Digital syndrome Craniofacial abnormality OFD1 Nephronophthisis NPHP1-5 Uncertain Basal body and cilia Retinitis pigmentosa RPGR Retinal transport DNAH11 Dynein arms For ciliopathies, see Afzelius, 2004 ; Badano et al., 2006 ; Fliegauf, 2007 The cell biological basis of ciliary disease Wallace F. The Journal of Cell Biology, Vol. 180, No. 1, January 14, –21

44
DNAH5 gén 79 exonból áll (kromoszóma:5p15-p14 régió).
H: nehéz lánc A: axonémális azaz csilló, DN: dinein Primary ciliary dyskinesia 3/ Kartagener syndrome 2 –eltérő allélok DNAI1 gén 20 exonból áll (kromoszóma :9-21-p13 régió) I. Intermedier lánc Primary ciliary dyskinesia 1/ Kartagener syndrome 1 DNAH11 gén 82 exonból áll (kromoszóma: 7p21 regió) Primary ciliary dyskinesia 7/ Kartagener Syndrome 3
 I. Intermedier lánc. Primary ciliary dyskinesia 1/ Kartagener syndrome 1. DNAH11 gén 82 exonból áll (kromoszóma: 7p21 regió) Primary ciliary dyskinesia 7/ Kartagener. Syndrome 3.")
45
Leggyakoribb tünetek: Bronchiectasis, infertilitás és situs inversus
Lokális, irreverzibilis dilatáció a bronchusokban, amely gyulladással jár. Gyakran nem megfelelő a szecernált váladék továbbhaladása ezen a területen. Gyakori a bakteriális infekció ezeken a területeken. Leggyakoribb tünetek: Bronchiectasis, infertilitás és situs inversus

46
Situs inversus Schematic illustration of normal left–right body asymmetry (situs solitus) and five laterality defects that affect the lungs, heart, liver, stomach and spleen. By their vigorous circular movements, motile monocilia at the embryonic node generate a leftward flow of extra-embryonic fluid (nodal flow). b | The nodal vesicular parcel (NVP) model predicts that vesicles filled with morphogens (such as sonic hedgehog and retinoic acid) are secreted from the right side of the embryonic node and transported to the left side by nodal flow, where they are smashed open by force27. The released contents probably bind to specific transmembrane receptors in the axonemal membrane of cilia on the left side. The consequent initiation of left-sided intracellular Ca2+ release induces downstream signalling events that break bilaterality. In this model, the flow of extra-embryonic fluid is not detected by cilia-based mechanosensation. c | In the two-cilia model, non-sensing motile cilia in the centre of the node create a leftward nodal flow that is mechanically sensed through passive bending of non-motile sensory cilia at the periphery of the node26. Bending of the cilia on the left side leads to a left-sided release of Ca2+ that initiates the establishment of body asymmetry. Embrió: mozgó csillók egy jobbról balra irányuló folyadékmozgást okoznak az embrióban (nodális flow). Ez egy morfogén grádienst hoz létre, ami felelős a jobb ill. bal oldali test asszimetriáért Hiba: situs inversust azaz eltérő asszimetriát okoz
 and five laterality defects that affect the lungs, heart, liver, stomach and spleen. By their vigorous circular movements, motile monocilia at the embryonic node generate a leftward flow of extra-embryonic fluid (nodal flow). b | The nodal vesicular parcel (NVP) model predicts that vesicles filled with morphogens (such as sonic hedgehog and retinoic acid) are secreted from the right side of the embryonic node and transported to the left side by nodal flow, where they are smashed open by force27. The released contents probably bind to specific transmembrane receptors in the axonemal membrane of cilia on the left side. The consequent initiation of left-sided intracellular Ca2+ release induces downstream signalling events that break bilaterality. In this model, the flow of extra-embryonic fluid is not detected by cilia-based mechanosensation. c | In the two-cilia model, non-sensing motile cilia in the centre of the node create a leftward nodal flow that is mechanically sensed through passive bending of non-motile sensory cilia at the periphery of the node26. Bending of the cilia on the left side leads to a left-sided release of Ca2+ that initiates the establishment of body asymmetry. Embrió: mozgó csillók egy jobbról balra irányuló folyadékmozgást okoznak az embrióban (nodális flow). Ez egy morfogén grádienst hoz létre, ami felelős a jobb ill. bal oldali test asszimetriáért. Hiba: situs inversust azaz eltérő asszimetriát okoz.")
47
Német elmegyógyász Alois Alzheimer írta le 1901-ben.
Alzheimer kór Német elmegyógyász Alois Alzheimer írta le ben. Dementia-val járó, gyógyíthatatlan neurodegenerációs betegség Átlagos életkor amikor kezdődnek a tünetek: 65 éves kor Legjellemzőbb tünetek: rövid távú memória vesztés ill. csökkenés kognitív képességek csökkenése agresszivitás, szétszórtság, hosszú távú memória vesztés Morfológiai változások Extracelluláris: Amyloid plakkok Intracelluláris: Neurofibrilláris aggregátumok

48
A tau fehérje és az Alzheimer kór kapcsolata
ha a tau fehérjék egymással összekapcsolódva aggregátumokat képeznek a mikrotubulusok dezintegrálódnak és a neuron funkcióképessége csökken ill. később a sejt elpusztul

49
Aggregált tau: PHF (paired helical filaments)
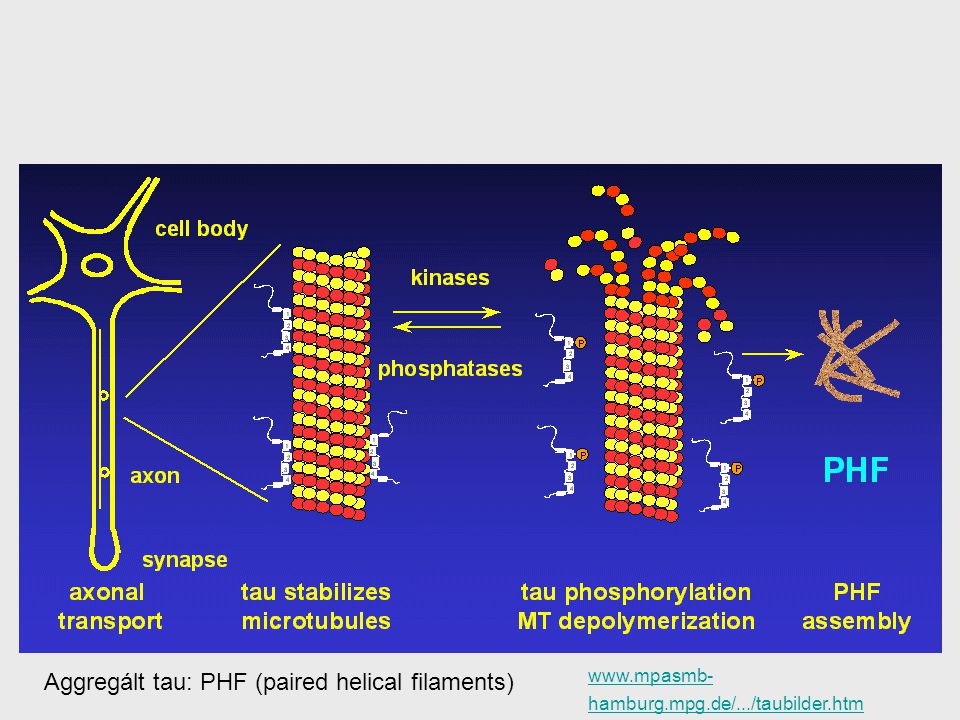
50
Miért és mikor okoz a tau fehérje neuron sejthalált?
Mi indítja be a hiperfoszforilációt? tau hiperfoszforiláció

51
A beta amyloid fehérjék keletkezése APP-ből
ICD:intracellular domain transzmembrán régió Beta-amyloid proteins cytosol Az APP fehérje, amelyből proteolítikus hasítás révén a sejttoxikus beta- amyloid fehérjék ( 40, 42) származnak oly módon, hogy az (alfa), béta, és gamma szekretáz enzimek hasítják a fehérjét
 származnak oly módon, hogy az. (alfa), béta, és gamma szekretáz enzimek hasítják a fehérjét.")
52
Alzheimer kór és a beta-amyloid fehérjék
Beta-amyloid plakk képződés az APP fehérje hasítása révén Ez indítja be az amiloid kaszkádot aminek eredményeként a sejtekben a tau fehérje hiperfoszforilálódik és aggregátumokat képez

53
A tau fehérjék hiperfoszforilációját a beta amyloid fehérjék okozhatják

Hasonló előadás







 prokariótában>")
 Minden fiú => x(x fiú …) Két prímszám=> x y( x prímszám y.>")



>")


 „késői” gének (late - L) 4.7 kb SV40 genom - kicsiny „tanulóvírus” fertőzést követően először.>")



