Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
NATURE NEWS BLOG G-protein-coupled receptors take chemistry Nobel
10 Oct 2012 Category: Chemistry The prize-winning receptor (blue), shown here bound to a signalling molecule (yellow) and activating a G-protein (red, gold and green). Brian K. Kobilka and Robert J. Lefkowitz have won this year’s chemistry Nobel for their work “crucial for understanding how G-protein–coupled receptors function”. Lefkowitz works at the Howard Hughes Medical Institute and Duke University in Durham, North Carolina. Kobilka is at Stanford University School of Medicine in California. The Nobel committee cites Lefkowitz’s work using radioactivity to trace cells’ receptors, which allowed researchers to start understanding how they work. Kobilka picked up the trail and started working to isolate the gene encoding one of the receptors earlier scrutinized by Lefkowitz. Then, in 2011, Kobilka’s team managed to tease out the structure of the one receptor actually in complex with its G-protein (see ‘Cell signalling caught in the act’). Read more about G-protein-coupled receptors in this collection of research and news published by Natureover the past few years.
, shown here bound to a signalling molecule (yellow) and activating a G-protein (red, gold and green). Brian K. Kobilka and Robert J. Lefkowitz have won this year’s chemistry Nobel for their work crucial for understanding how G-protein–coupled receptors function . Lefkowitz works at the Howard Hughes Medical Institute and Duke University in Durham, North Carolina. Kobilka is at Stanford University School of Medicine in California. The Nobel committee cites Lefkowitz’s work using radioactivity to trace cells’ receptors, which allowed researchers to start understanding how they work. Kobilka picked up the trail and started working to isolate the gene encoding one of the receptors earlier scrutinized by Lefkowitz. Then, in 2011, Kobilka’s team managed to tease out the structure of the one receptor actually in complex with its G-protein (see ‘Cell signalling caught in the act’). Read more about G-protein-coupled receptors in this collection of research and news published by Natureover the past few years.")
2
Jelátadó komplexek és a jelpályák kialakulása
Sejten belüli jelátadó molekulák szerkezeti egységei, az enzimhez kapcsolt receptorok jelátadási mechanizmusa I Jelátadó komplexek és a jelpályák kialakulása A ligandumnak megfelelő, specifikus receptorhoz való kötődést a jetovábbításban résztvevő fehérjék sorának kötődése követi az aktivált receptor közelébe, ennek következtében kialakul a jelátadó komplex. A jelátadó komplex elemei szerkezeti egységeik segítségével kapcsolódnak egymáshoz

3
A jelátadás mechanizmusa
=> a jelátadó komplex kialakulása, aktiválódása megváltoztatja a jelátadási kaszkádban résztvevő effektor fehérjék szerkezetét, elhelyezkedését, vagy funkcióját A jelátadási kaszkád (jelpálya) a jelátadási hálózat része => az információ továbbítást biztosítja A jelátadási hálózat biztosítja, hogy a sejtek összehangolt választ adjanak a környezet sokféle jelére
 a jelátadási hálózat része. => az információ továbbítást biztosítja. A jelátadási hálózat biztosítja, hogy a sejtek összehangolt választ adjanak a környezet sokféle jelére.")
4
Példa: T sejtek jelátviteli hálózata

5
Példa: TLR jelátvitel

6
Jeltadó (signaling) komplexek kialakulása
A jeltadó komplexek kialakulása protein-protein vagy protein-lipid kölcsönhatásokon alapul, amelyeket protein domének és speciális szekvencia motívumok kapcsolódása közvetít Szekvencia motívumok: pYXXM Pro-rich PIP2 ---Pro-rich/ P-Ser ----NPEY----PI3P | | | | | | SH SH3 PH WW PTB FYVE Protein domének Példák: -Src-homology-2 (SH2) doméneket tartalmazó fehérjék a P-Tyr aminosavmaradékot tartalmazó motívummal kapcsolódnak (pl.YMXM) -pleksztrin homológ (PH) doméneket tartalmazó fehérjék foszfolipidekhez, pl. PI-3,4,5P-hoz kapcsolódnak Ezek a szerkezeti egységek (domének) funkcionálisan önálló egységek és a jelátadó molekulákban egymástól függetlenül, különböző számban fordulnak elő
 doméneket tartalmazó fehérjék a P-Tyr aminosavmaradékot tartalmazó motívummal kapcsolódnak (pl.YMXM) -pleksztrin homológ (PH) doméneket tartalmazó fehérjék foszfolipidekhez, pl. PI-3,4,5P-hoz kapcsolódnak. Ezek a szerkezeti egységek (domének) funkcionálisan önálló egységek és a jelátadó molekulákban egymástól függetlenül, különböző számban fordulnak elő.")
7
A jeltovábbító fehérjék moduláris szerkezete
Modular nature of signaling proteins. Representative members from various SH2 domain families and the positional organization of the domain modules within these proteins are illustrated. See the text for further details. Additional information on individual domains can be found at and Abbreviations: CH, calponin homology; C1, PKC conserved region 1 phorbol ester binding; C2, PKC conserved region 2 calcium-dependent phospholipid binding; DNA BD, DNA-binding domain; EF, EF-hand; FCH, Fes/CIP4 homology domain; 4H, four-helix bundle; GAP, GTPase-activating domain; GEF, GDP¯GTP exchange factor; PH, pleckstrin homology domain; PLC, phospholipase C; PTB, phosphotyrosine-binding domain; PTPc/DSPc, protein tyrosine phosphatase, unknown specificity; SAM, sterile alpha motif; SH2, Src-homology 2 domain; SH3, Src-homology 3 domain; SOCS, suppressor of cytokine signaling; STAT signal transducer and activator of transcription; TA, transcriptional activation domain; Uba, ubiquitin-associated domain.

8
A receptor tirozin kinázok jelátvitelében szereplő protein modulok (a) és dokkoló fehérjék (b)
a) A moduláris jelátadó fehérjék tartalmaznak SH2 és PTB (foszfotirozin kötő) PH, SH3, WW, PDZ FYVE doméneket b) Horgonyzó (dokkoló) fehérjék Protein Modules and Docking Proteins that Participate in Signaling via Receptor Tyrosine Kinases (A) Protein modules implicated in the control of intracellular signaling pathways. Tyrosine phosphorylated, activated RTKs form a complex with SH2 and PTB domains of signaling proteins. SH2 domains bind to pTyr sites in activated receptors while PTB domains bind to tyrosine phosphorylated and nonphosphorylated regions in RTKs. PH domains bind to different phosphoinositides leading to membrane association. SH3 and WW domains bind to proline-rich sequences in target proteins. PDZ domains bind to hydrophobic residues at the C termini of target proteins. FYVE domains bind specifically to PdtIns(3)P. While adaptor proteins such as Grb2 or Nck contain only SH2 and SH3 domains, other signaling proteins contain additional enzymatic activities such as protein kinases (Src,PKB), PTPase (Shp2) phospholipase C (PLC), Ras-GAP or Rho-GRF (Vav). (B) Docking proteins that function as platforms for recruitment of signaling proteins. All docking proteins contain a membrane targeting region in their N termini. FRS2 is targeted to the membrane by myristoylation, and LAT is targeted to the cell membrane by a transmembrane domains (TM) and by palmytoylation. Most docking proteins are targeted to the cell membrane by their PH domains. Docking proteins contain multiple pTyr phosphorylation sites that function as binding sites for SH2 domains of a variety of signaling proteins. PDZ is an acronym combining the first letters of three proteins — post synaptic density protein (PSD95), Drosophila disc large tumor suppressor (DlgA), and zonula occuldens-1 protein (zo-1) — which were first discovered to share the domain. These domains help anchor transmembrane proteins to the cytoskeleton and hold together signaling complexes
 A moduláris jelátadó fehérjék tartalmaznak. SH2 és PTB (foszfotirozin kötő) PH, SH3, WW, PDZ FYVE doméneket. b) Horgonyzó (dokkoló) fehérjék. Protein Modules and Docking Proteins that Participate in Signaling via Receptor Tyrosine Kinases. (A) Protein modules implicated in the control of intracellular signaling pathways. Tyrosine phosphorylated, activated. RTKs form a complex with SH2 and PTB domains of signaling proteins. SH2 domains bind to pTyr sites in activated. receptors while PTB domains bind to tyrosine phosphorylated and nonphosphorylated regions in RTKs. PH domains. bind to different phosphoinositides leading to membrane association. SH3 and WW domains bind to proline-rich. sequences in target proteins. PDZ domains bind to hydrophobic residues at the C termini of target proteins. FYVE. domains bind specifically to PdtIns(3)P. While adaptor proteins such as Grb2 or Nck contain only SH2 and SH3 domains, other signaling proteins contain additional enzymatic activities such as protein kinases (Src,PKB), PTPase (Shp2) phospholipase C (PLC), Ras-GAP or Rho-GRF (Vav). (B) Docking proteins that function as platforms for recruitment of signaling proteins. All docking proteins contain a. membrane targeting region in their N termini. FRS2 is targeted to the membrane by myristoylation, and LAT is. targeted to the cell membrane by a transmembrane domains (TM) and by palmytoylation. Most docking proteins are. targeted to the cell membrane by their PH domains. Docking proteins contain multiple pTyr phosphorylation sites that. function as binding sites for SH2 domains of a variety of signaling proteins. PDZ is an acronym combining the first letters of three proteins — post synaptic density protein (PSD95), Drosophila disc large tumor suppressor (DlgA), and zonula occuldens-1 protein (zo-1) — which were first discovered to share the domain. These domains help anchor transmembrane proteins to the cytoskeleton and hold together signaling complexes.")
9
Az SH2 domének által közvetítet protein-protein kölcsönhatás
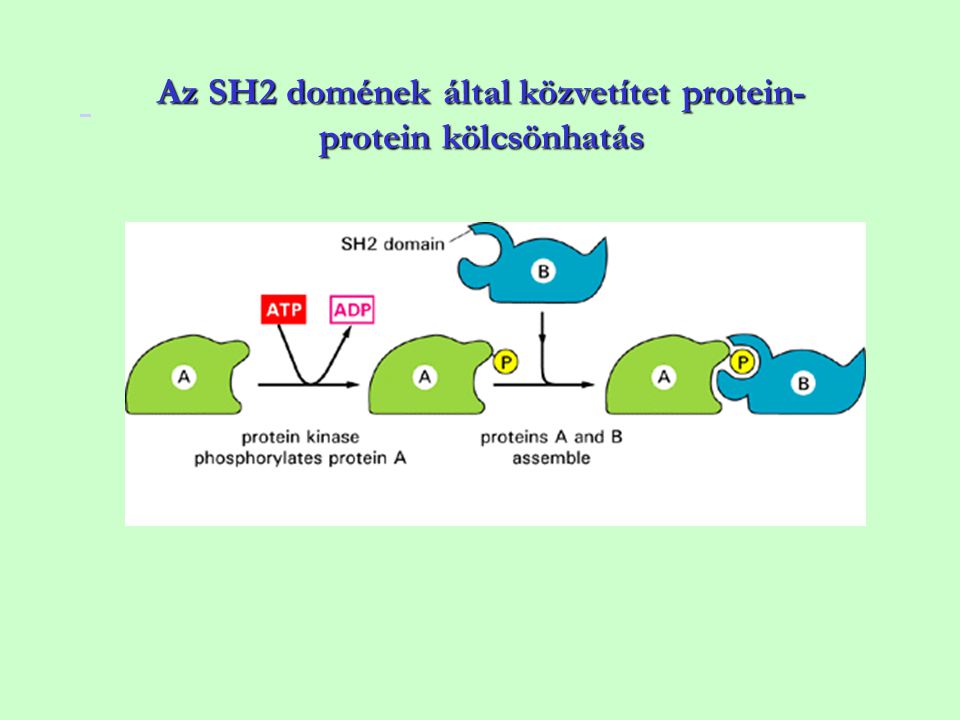
10
A pYEEI peptid- ligandumot kötő v-src SH2 doménje
Az SH2 domének ligandum specificitása etérő, mutációval megváltoztatható Thr Trp SH2 domains contain a central anti-parallel b-sheet surrounded by two a-helices. The phosphopeptide generally binds as an extended b-strand that lies at right angles to the SH2 b-sheet. Conserved residues contribute to the hydrophobic core or are involved in pY recognition while more variable residues contribute to specific recognition of C-terminal residues. An invariant Arg residue in the SH2 domain coordinates the phosphate oxygens of pY and is essential for high affinity phosphopeptide binding. The figure shows the SH2 domain of v-src bound to a pYEEI peptide ligand. Structural analysis provides a basis for understanding SH2 domain binding specificity. Crystal structures of the Grb2 and Src SH2 domains bound to optimal phosphopeptides illustrate the different modes of ligand binding. A switch in the ligand preference of the Src SH2 domain to pTyr-Val-Asn-Val is achieved through mutation of a threonine residue, highlighted in blue on the surface of the Src SH2, to the tryptophan shown as red in the Src T215W SH2 domain. In a similar fashion, the molecular architecture of the Grb2 SH2 domain uses a tryptophan (red) to define its binding to pTyr-X-Asn ligands. (For more details see Pawson, T., Gish, G., and Nash, P. Trends in Cell Biology Vol.11 No.12 December 2001) Centrális anti paralell b lemez + két alfa helix, kb.100 aa Konzervatív Arg – tyr foszfát oxigénje közötti kapcsolat – elengedhetetlen a nagy affinitású kötéshez Kd= nM
 to define its binding to pTyr-X-Asn ligands. (For more details see Pawson, T., Gish, G., and Nash, P. Trends in Cell Biology Vol.11 No.12 December 2001) Centrális anti paralell b lemez + két alfa helix, kb.100 aa. Konzervatív Arg – tyr foszfát oxigénje közötti kapcsolat – elengedhetetlen a nagy affinitású kötéshez Kd= nM.")
11
Src, Fyn, Yes, Fgr, Lyn, Hck, Lck, Blk, and Yrk
SH2 Domains [Crit. Rev. Oncol, Hematol. 40, , 2001] [Trends Cell Biol. 11, , 2001] - Src- az először leírt citoplazmában levő protein tirozin kináz (PTK), onkogén termék Az Src PTK család: Src, Fyn, Yes, Fgr, Lyn, Hck, Lck, Blk, and Yrk [BBA, 1287, , 1996] [BBA 1422(2): , 1999] Src Homology Region 2 (SH2) – először itt írták le ~100 aminosav, a katalitikus doméntől (SH1) eltérő szekvencia
, onkogén termék. Az Src PTK család: Src, Fyn, Yes, Fgr, Lyn, Hck, Lck, Blk, and Yrk. [BBA, 1287, , 1996] [BBA 1422(2): , 1999] Src Homology Region 2 (SH2) – először itt írták le. ~100 aminosav, a katalitikus doméntől (SH1) eltérő szekvencia.")
12
Az SH2 domén tirozinon foszforilált fehérjéket köt:
p-Y-x-x-x motívumot specifitás: pY- hidrofil-hidrofil-hidrofób ( src, lck) pY-hidrofób – X – hidrofób (Shp-2, Plcg) Szerkezet: antiparalel beta lemez, + 2 alfa helix alkotja Kötődés függ: lokális szubsztrát koncentrációtól, más domének szabályozó hatásától
 pY-hidrofób – X – hidrofób (Shp-2, Plcg) Szerkezet: antiparalel beta lemez, + 2 alfa helix alkotja. Kötődés függ: lokális szubsztrát koncentrációtól, más domének szabályozó hatásától.")
13
SH2 domént tartalmazó jelátadó molekulák kötődése az aktivált PDGF receptorhoz
Figure The binding of SH2-containing intracellular signaling proteins to an activated PDGF receptor. (A) This drawing of a PDGF receptor shows five of the tyrosine autophosphorylation sites, three in the kinase insert region and two on the C-terminal tail, to which the three signaling proteins shown bind as indicated. The numbers on the right indicate the positions of the tyrosines in the polypeptide chain. These binding sites have been identified by using recombinant DNA technology to mutate specific tyrosines in the receptor. Mutation of tyrosines 1009 and 1021, for example, prevents the binding and activation of PLC-γ, so that receptor activation no longer stimulates the inositol phospholipid signaling pathway. The locations of the SH2 (red) and SH3 (blue) domains in the three signaling proteins are indicated. (Additional autophosphorylation sites on this receptor are not shown, including those that serve as binding sites for the cytoplasmic tyrosine kinase Src and the adaptor proteins Grb2 and Shc, discussed later.) (B) The three-dimensional structure of an SH2 domain, as determined by x-ray crystallography. The binding pocket for phosphotyrosine is shown in yellow on the right, and a pocket for binding a specific amino acid side chain (isoleucine, in this case) is shown in yellow on the left (see also Figure 3-40). (C) The SH2 domain is a compact, “plug-in” module, which can be inserted almost anywhere in a protein without disturbing the protein's folding or function (see Figure 3-19). Because each domain has distinct sites for recognizing phosphotyrosine and for recognizing a particular amino acid side chain, different SH2 domains recognize phosphotyrosine in the context of different flanking amino acid sequences. (B, based on data from G. Waksman et al., Cell 72:1–20, © Elsevier.) 3-6 aminosav a C terminális felé, meghatározza a kötődés specifitását
 This drawing of a PDGF receptor shows five of the tyrosine autophosphorylation sites, three in the kinase insert region and two on the C-terminal tail, to which the three signaling proteins shown bind as indicated. The numbers on the right indicate the positions of the tyrosines in the polypeptide chain. These binding sites have been identified by using recombinant DNA technology to mutate specific tyrosines in the receptor. Mutation of tyrosines 1009 and 1021, for example, prevents the binding and activation of PLC-γ, so that receptor activation no longer stimulates the inositol phospholipid signaling pathway. The locations of the SH2 (red) and SH3 (blue) domains in the three signaling proteins are indicated. (Additional autophosphorylation sites on this receptor are not shown, including those that serve as binding sites for the cytoplasmic tyrosine kinase Src and the adaptor proteins Grb2 and Shc, discussed later.) (B) The three-dimensional structure of an SH2 domain, as determined by x-ray crystallography. The binding pocket for phosphotyrosine is shown in yellow on the right, and a pocket for binding a specific amino acid side chain (isoleucine, in this case) is shown in yellow on the left (see also Figure 3-40). (C) The SH2 domain is a compact, plug-in module, which can be inserted almost anywhere in a protein without disturbing the protein s folding or function (see Figure 3-19). Because each domain has distinct sites for recognizing phosphotyrosine and for recognizing a particular amino acid side chain, different SH2 domains recognize phosphotyrosine in the context of different flanking amino acid sequences. (B, based on data from G. Waksman et al., Cell 72:1–20, © Elsevier.) 3-6 aminosav a C terminális felé, meghatározza a kötődés specifitását.")
14
Peptid kötődése az SH2 doménhez
SH2 domén - modell Figure The evolutionary trace method applied to the SH2 domain. (A) Front and back views of a space-filling model of the SH2 domain, with evolutionarily conserved amino acids on the protein surface colored yellow, and those more toward the protein interior colored red. (B) The structure of the SH2 domain with its bound polypeptide. Here, those amino acids located within 0.4 nm of the bound ligand are colored blue. The two key amino acids of the ligand are yellow, and the others are purple. (Adapted from O. Lichtarge, H.R. Bourne, and F.E. Cohen, J. Mol. Biol. 257:342–358, 1996.) Peptid kötődése az SH2 doménhez Az evolúció során konzervált aminosavak
 Front and back views of a space-filling model of the SH2 domain, with evolutionarily conserved amino acids on the protein surface colored yellow, and those more toward the protein interior colored red. (B) The structure of the SH2 domain with its bound polypeptide. Here, those amino acids located within 0.4 nm of the bound ligand are colored blue. The two key amino acids of the ligand are yellow, and the others are purple. (Adapted from O. Lichtarge, H.R. Bourne, and F.E. Cohen, J. Mol. Biol. 257:342–358, 1996.) Peptid kötődése az SH2 doménhez. Az evolúció során konzervált aminosavak.")
15
KÖTÉS: p-Y: -mély zseb, pozitív töltés, konzervatív Arg peptid: eltérő, hidrofil, hidrofób kölcsönhatások oldalláncok hossza Kötés két lépésben: gyenge asszociációt erős kötődés követi KD=<nM nagy affinitású kötés FUNKCIÓ: Sejten belüli lokalizáció megváltoztatása Konformáció változás Jel sokszorozás

16
Mi az SH2 domén funkciója: -enzim-szubsztrát közelség
-Konformáció változás enzim aktiválódás -Tyr foszforiláció

17
SH3 domén Két SH3 domén osztály: RKXXPXXP és PXXPXR Kd = mM tartomány
PPPVPPRRR (SOS) 5 antiparalel b szál 2 b lemezt alkot, ligandum kötő hely: konzervált aromás aminosavak (hidrofób) + 2 töltéssel rendelkező, variábilis hurok Két SH3 domén osztály: RKXXPXXP és PXXPXR Kd = mM tartomány The basic fold of SH3 domains contains five anti-parallel beta-strands packed to form two perpendicular beta-sheets. The ligand-binding site consists of a hydrophobic patch that contains a cluster of conserved aromatic residues and is surrounded by two charged and variable loops. The figure shows a Sem5 C-terminal SH3 domain complexed to the mSos-derived sequence PPPVPPRRR.
 5 antiparalel b szál 2 b lemezt alkot, ligandum kötő hely: konzervált aromás aminosavak (hidrofób) + 2 töltéssel rendelkező, variábilis hurok. Két SH3 domén osztály: RKXXPXXP és PXXPXR. Kd = mM tartomány. The basic fold of SH3 domains contains five anti-parallel beta-strands packed to form two perpendicular beta-sheets. The ligand-binding site consists of a hydrophobic patch that contains a cluster of conserved aromatic residues and is surrounded by two charged and variable loops. The figure shows a Sem5 C-terminal SH3 domain complexed to the mSos-derived sequence PPPVPPRRR.")
18
SH3 domének [Cell Signal. 11(4): , 1999., Cell Sci. 114 (7) ] kb. 60 aminosav, fehérje-fehérje kölcsönhatásokat segíti elő SH2 doménhez hasonlóan enzimekben, szerkezeti fehérjékben, kis adapter fehérjékben fordul elő Sok jeláadó molekula rendelkezik mind SH2, mind SH3 doménnel –Az SH2-től eltérően, SH3 domének már az élesztőben megjelennek ==> SH3 domének az evolúció során korán megjelenő, primitív funkcióval rendelkeznek A kötődési motívum: prolinban gazdag PXXPPP (hidrofób) XP [Science, 259, , 1993] SH3 domént tartalmazó fehérjék sora tartozik a citoszkeletonnal kölcsönhatásba lépő molekulák közé – miozin, spektrin, fodrin- a sejt struktúra és mozgásképesség kialakításában fontos
![SH3 domének [Cell Signal. 11(4): , 1999., Cell Sci. 114 (7) ] kb. 60 aminosav, fehérje-fehérje kölcsönhatásokat segíti elő.](http://slideplayer.hu/slide/2091063/8/images/18/SH3+dom%C3%A9nek+%5BCell+Signal.+11%284%29%3A+%2C+1999.%2C+Cell+Sci.+114+%287%29+%5D+kb.+60+aminosav%2C+feh%C3%A9rje-feh%C3%A9rje+k%C3%B6lcs%C3%B6nhat%C3%A1sokat+seg%C3%ADti+el%C5%91..jpg "SH2 doménhez hasonlóan enzimekben, szerkezeti fehérjékben, kis adapter fehérjékben fordul elő. Sok jeláadó molekula rendelkezik mind SH2, mind SH3 doménnel –Az SH2-től eltérően, SH3 domének már az élesztőben megjelennek. ==> SH3 domének az evolúció során korán megjelenő, primitív funkcióval rendelkeznek. A kötődési motívum: prolinban gazdag. PXXPPP (hidrofób) XP. [Science, 259, , 1993] SH3 domént tartalmazó fehérjék sora tartozik a citoszkeletonnal kölcsönhatásba lépő molekulák közé – miozin, spektrin, fodrin- a sejt struktúra és mozgásképesség kialakításában fontos.")
19
SH3 domén kölcsönhatása a ligandummal

20
Az Src kináz autoregulációja és aktiválódása
Kináz aktivitás nő: Molekulán belüli kölcsönhatások:

21
Rous sarcoma virus: C terminális 18 aa hiányzik – konstitutívan aktív
Az Src aktivitás szabályozása Regulation of Src activity and its activation by an oncogenic mutation. (a) Domain structure of c-Src and v-Src. Phosphorylation of tyrosine 527 by Csk, another cellular tyrosine kinase, inactivates the Src kinase activity. The transforming v-Src oncoprotein encoded by Rous sarcoma virus is missing the C-terminal 18 amino acids including tyrosine 527 and thus is constitutively active. (b) Effect of phosphorylation on c-Src conformation. Binding of phosphotyrosine 527 to the SH2 domain induces conformational strains in the SH3 and kinase domains, distorting the kinase active site so it is catalytically inactive. The kinase activity of c-Src is normally activated by removing the phosphate on tyrosine 527. [Adapted from T. Pawson, 1997, Nature 385:582. See also W. Xu et al., 1997, Nature 385:595; and F. Sichrei et al., 1997, Nature 385:602.] Regulation of Src activity and its activation by an oncogenic mutation. (a) Domain structure of c-Src and v-Src. Phosphorylation of tyrosine 527 by Csk, another cellular tyrosine kinase, inactivates the Src kinase activity. The transforming v-Src oncoprotein encoded by Rous sarcoma virus is missing the C-terminal 18 amino acids including tyrosine 527 and thus is constitutively active. (b) Effect of phosphorylation on c-Src conformation. Binding of phosphotyrosine 527 to the SH2 domain induces conformational strains in the SH3 and kinase domains, distorting the kinase active site so it is catalytically inactive. The kinase activity of c-Src is normally activated by removing the phosphate on tyrosine 527. [Adapted from T. Pawson, 1997, Nature 385:582. See also W. Xu et al., 1997, Nature 385:595; and F. Sichrei et al., 1997, Nature 385:602.]
 Domain structure of c-Src and v-Src. Phosphorylation of tyrosine 527 by Csk, another cellular tyrosine kinase, inactivates the Src kinase activity. The transforming v-Src oncoprotein encoded by Rous sarcoma virus is missing the C-terminal 18 amino acids including tyrosine 527 and thus is constitutively active. (b) Effect of phosphorylation on c-Src conformation. Binding of phosphotyrosine 527 to the SH2 domain induces conformational strains in the SH3 and kinase domains, distorting the kinase active site so it is catalytically inactive. The kinase activity of c-Src is normally activated by removing the phosphate on tyrosine 527. [Adapted from T. Pawson, 1997, Nature 385:582. See also W. Xu et al., 1997, Nature 385:595; and F. Sichrei et al., 1997, Nature 385:602.] Regulation of Src activity and its activation by an oncogenic mutation. (a) Domain structure of c-Src and v-Src. Phosphorylation of tyrosine 527 by Csk, another cellular tyrosine kinase, inactivates the Src kinase activity. The transforming v-Src oncoprotein encoded by Rous sarcoma virus is missing the C-terminal 18 amino acids including tyrosine 527 and thus is constitutively active. (b) Effect of phosphorylation on c-Src conformation. Binding of phosphotyrosine 527 to the SH2 domain induces conformational strains in the SH3 and kinase domains, distorting the kinase active site so it is catalytically inactive. The kinase activity of c-Src is normally activated by removing the phosphate on tyrosine 527. [Adapted from T. Pawson, 1997, Nature 385:582. See also W. Xu et al., 1997, Nature 385:595; and F. Sichrei et al., 1997, Nature 385:602.]")
22
SH2 és SH3 doméneket tartalmazó fehérjék a p-tyr horgonyzó
helyet összekapcsolják különféle jeltovábbító rendszerekkel Grb2: Adapter fehérje Az evolúció során konzervált Sokrétű funkció

23
PH (pleksztrin homológ) domén
PLC PIP3 Két merőleges anti-paralell b lemez + C terminális a amfipatikus helix, a kötésben bázikus aminosavak vesznek részt Despite unusually divergent primary sequences, PH domains share a conserved fold made up of a b-barrel composed of two roughly perpendicular, anti-parallel beta-sheets and a C-terminal alpha amphipathic -helix. PH domains bind to their inositol phosphate ligands via a binding surface composed primarily of residues from the beta1/beta2, beta3/beta4, and beta6/beta7 loops. Basic residues in the beta1-beta2 loop are especially important for binding the phosphatidylinositide phosphates, by establsihing a positive electric potential on the face of the PH domain. Comparison of different PH domains with distinct phosphoinosite binding specificities has revealed insights into what structural features dictate binding specificity. For example, a tyrosine residue located in the beta-strand is strictly conserved in PH domains that bind PI3- kinase products with high affinity but not in other PH domains. Interestingly, PH domains have a strikingly similar structure to PTB domains, the Enabled / Vasp homology (EVH1/WH1) and the Ran-binding domain (RanBD) of RBP2, despite the absence of any sequence homology. The figure shows the complex between the PH domain of Phospholipase C-d and inositol-(1,4,5)-trisphosphate (red). Despite unusually divergent primary sequences, PH domains share a conserved fold made up of a b-barrel composed of two roughly perpendicular, anti-parallel beta-sheets and a C-terminal alpha amphipathic -helix. PH domains bind to their inositol phosphate ligands via a binding surface composed primarily of residues from the beta1/beta2, beta3/beta4, and beta6/beta7 loops. Basic residues in the beta1-beta2 loop are especially important for binding the phosphatidylinositide phosphates, by establsihing a positive electric potential on the face of the PH domain. Comparison of different PH domains with distinct phosphoinosite binding specificities has revealed insights into what structural features dictate binding specificity. For example, a tyrosine residue located in the beta-strand is strictly conserved in PH domains that bind PI3- kinase products with high affinity but not in other PH domains. Interestingly, PH domains have a strikingly similar structure to PTB domains, the Enabled / Vasp homology (EVH1/WH1) and the Ran-binding domain (RanBD) of RBP2, despite the absence of any sequence homology. The figure shows the complex between the PH domain of Phospholipase C-d and inositol-(1,4,5)-trisphosphate (red).
 and the Ran-binding domain (RanBD) of RBP2, despite the absence of any sequence homology. The figure shows the complex between the PH domain of Phospholipase C-d and inositol-(1,4,5)-trisphosphate (red). Despite unusually divergent primary sequences, PH domains share a conserved fold made up of a b-barrel composed of two roughly perpendicular, anti-parallel beta-sheets and a C-terminal alpha amphipathic -helix. PH domains bind to their inositol phosphate ligands via a binding surface composed primarily of residues from the beta1/beta2, beta3/beta4, and beta6/beta7 loops. Basic residues in the beta1-beta2 loop are especially important for binding the phosphatidylinositide phosphates, by establsihing a positive electric potential on the face of the PH domain. Comparison of different PH domains with distinct phosphoinosite binding specificities has revealed insights into what structural features dictate binding specificity. For example, a tyrosine residue located in the beta-strand is strictly conserved in PH domains that bind PI3- kinase products with high affinity but not in other PH domains. Interestingly, PH domains have a strikingly similar structure to PTB domains, the Enabled / Vasp homology (EVH1/WH1) and the Ran-binding domain (RanBD) of RBP2, despite the absence of any sequence homology. The figure shows the complex between the PH domain of Phospholipase C-d and inositol-(1,4,5)-trisphosphate (red).")
24
PLEKSZTRIN homológ domén (PH)
A PH domén szerkezete A PH domének az SH2 és SH3 doménekkel összehasonlítva kevésbé egységes szerkezetűek. Az aminosav sorrend eltéréseinek ellenére a PH domének három dimenziós szerkezete nagy mértékű azonosságot mutat. A szerkezet hat b lánc által alkotott, félig nyitott hordó, melyet egy a helix fed le A PKC fő szubsztrátja trombocitákban a pleksztrin, melynek C és N terminális doménje kb. 100 aminosavból álló, hasonló szerkezetet mutat. 100 körül van azoknak a fehérjéknek a száma amelyek a jeátvitelben szerepet játszanak vagy a citoszkeletonhoz kapcsolódnak és ehhez hasonló szerkezetet mutatnak. Ezeket a szerkezeti egységeket pleksztrin-homológ (PH) doméneknek nevezzük.
 doméneknek nevezzük.")
25
Az SH2 és SH3 doménekhez hasonlóan a PH domének nem foglalnak el egy specifikus helyet az adott fehérjében – funkcionálisan önállóak. Mi a liganduma a PH doménnek? A PH domének és a lipid kötő fehérjék szerkezeti hasonlósága arra utalt, hogy a PH domének lipofil molekulákat kötnek. -Kimutatták foszfoinozitidekhez való kötődésüket. -A kötésben a domén alján elhelyezkedő bázikus aminosavak csoportja vesz részt. - GST-PH fúziós fehérjék kölcsönhatásba kerülnek G fehérjékkel is - A „kiterjesztett” PH domén (PH + a környezete) kötődik lipidekhez (PIP2, PIP3) az N terminális részén keresztül, és G fehérjékhez is, a C terminális rész közvetítésével. ==> A PH domén kooperál a vele szomszédos doménekkel a fehérje-fehérje kölcsönhatások kialakításához
 kötődik lipidekhez (PIP2, PIP3) az N terminális részén keresztül, és G fehérjékhez is, a C terminális rész közvetítésével. ==> A PH domén kooperál a vele szomszédos doménekkel a fehérje-fehérje kölcsönhatások kialakításához.")
26
PÉLDÁK: PH domain Protein: Specifikus Foszfoinozitid Ligandum Phospholipase C ; mSos1 PI-4,5-P2 Btk Tyr Kinase; Grp1 PI-3,4,5-P3 Akt/PKB Ser/Thr Kinase PI-3,4-P2

27
A foszfatidil inozitol inozitol csoportja három helyen reverzibilisen foszforilálódhat –sokrétű funkció

28
-Kinázok -Foszfatázok -Lipázok

29
A PH domén funkciója: a fehérjék membránhoz kötése
kooperativitást igényel (beta- adrenergic receptor kinase )
")
30
PTB (phosphotyrosine binding) domén - Shc
HIIENPQpYFS Kötődő konszenzus szekvencia: NPXpY The PTB domains of Shc and IRS-1 contain two orthogonal b-sheets and connecting loops, and have very similar folds despite their low sequence similarity. Both have a C-terminal amphipathic a-helix capping one end of the b-sandwich. The N-terminal residues of the peptide ligand form an additional anti-parallel b-strand to the second b-sheet. The figure shows the PTB domain of Shc complexed to a HIIENPQpYFS peptide Phosphotyrosine binding (PTB) domains are residue modules that commonly bind Asn-Pro-X-Tyr motifs. The PTB domains of the docking proteins Shc and IRS-1 require ligand phosphorylation on the tyrosine residue (NPXpY) for binding. More N-terminal sequences are also required for high affinity binding and conferring specificity. The peptide binds as a b-strand to an anti-parallel b-sheet, while the NPXpY motif makes a turn, positioning the pY for recognition by basic residues. The PTB domains of proteins such as X11, Dab, Fe65 and Numb apparently recognize NPXY or related peptide motifs, but are not dependent on ligand phosphorylation. In addition, the Numb PTB domain can bind an unrelated peptide that forms a helical turn. The PTB domains of Shc and IRS-1 contain two orthogonal b-sheets and connecting loops, and have very similar folds despite their low sequence similarity. Both have a C-terminal amphipathic a-helix capping one end of the b-sandwich. The N-terminal residues of the peptide ligand form an additional anti-parallel b-strand to the second b-sheet. The figure shows the PTB domain of Shc complexed to a HIIENPQpYFS peptide
 domains are residue modules that commonly bind Asn-Pro-X-Tyr motifs. The PTB domains of the docking proteins Shc and IRS-1 require ligand phosphorylation on the tyrosine residue (NPXpY) for binding. More N-terminal sequences are also required for high affinity binding and conferring specificity. The peptide binds as a b-strand to an anti-parallel b-sheet, while the NPXpY motif makes a turn, positioning the pY for recognition by basic residues. The PTB domains of proteins such as X11, Dab, Fe65 and Numb apparently recognize NPXY or related peptide motifs, but are not dependent on ligand phosphorylation. In addition, the Numb PTB domain can bind an unrelated peptide that forms a helical turn. The PTB domains of Shc and IRS-1 contain two orthogonal b-sheets and connecting loops, and have very similar folds despite their low sequence similarity. Both have a C-terminal amphipathic a-helix capping one end of the b-sandwich. The N-terminal residues of the peptide ligand form an additional anti-parallel b-strand to the second b-sheet. The figure shows the PTB domain of Shc complexed to a HIIENPQpYFS peptide.")
31
PhosphoTyrosine-Binding (PTB) domén
A Shc fehérje 186 aminosavat tartalmazó szegmentjeként azonosították amely specifikusan kötődik foszfotirozin tartalmú fehérjékhez. [Science, 268, , 1995] [Nature, 378, , 1995] [J. Biol. Chem., 271, , 1996] [J. Biol. Chem., 271, , 1996] [Cell, 85, , 1996] [MCB, 17 (3), , 1997]
, , 1997]")
32
SH2 és PTB domének struktúrálisan alapvetően különbözőek
NPXpY SH2 és PTB domének struktúrálisan alapvetően különbözőek PTB nemcsak foszforilált tirozint köt. (pl.NPXY az IRS fehérjében)
")
33
Hasonlóság PH doménnel De ligandum más— Eltérő funkció
Structure of the IRS-1 targeting domain. (A) Ribbon diagram of the PH-PTB structure, with b-sheets shaded green, a-helices in turquoise, 310 turns colored indigo, and intervening coils or loops in brown. Residues within the PTB domain binding site (facing away) and putative PH domain binding site are colored dark gray and numbered; residues at the interface between the two domains are colored according to percent buried: red, %; orange, 25-50%; and yellow, 1-25%. (B) The PH/PTB domain interface viewed as an open book. The PH and PTB domains each were rotated 90o, relative to their orientations in A, but in opposite directions to expose the buried surface between them. Elements of secondary structure and contact residues are labeled. [PNAS 96, , 1999] Structure of the IRS-1 targeting domain. (A) Ribbon diagram of the PH-PTB structure, with b-sheets shaded green, a-helices in turquoise, 310 turns colored indigo, and intervening coils or loops in brown. Residues within the PTB domain binding site (facing away) and putative PH domain binding site are colored dark gray and numbered; residues at the interface between the two domains are colored according to percent buried: red, %; orange, 25-50%; and yellow, 1-25%. (B) The PH/PTB domain interface viewed as an open book. The PH and PTB domains each were rotated 90o, relative to their orientations in A, but in opposite directions to expose the buried surface between them. Elements of secondary structure and contact residues are labeled. [PNAS 96, , 1999]
 Ribbon diagram of the PH-PTB structure, with b-sheets shaded green, a-helices in turquoise, 310 turns colored indigo, and intervening coils or loops in brown. Residues within the PTB domain binding site (facing away) and putative PH domain binding site are colored dark gray and numbered; residues at the interface between the two domains are colored according to percent buried: red, %; orange, 25-50%; and yellow, 1-25%. (B) The PH/PTB domain interface viewed as an open book. The PH and PTB domains each were rotated 90o, relative to their orientations in A, but in opposite directions to expose the buried surface between them. Elements of secondary structure and contact residues are labeled. [PNAS 96, , 1999] Structure of the IRS-1 targeting domain. (A) Ribbon diagram of the PH-PTB structure, with b-sheets shaded green, a-helices in turquoise, 310 turns colored indigo, and intervening coils or loops in brown. Residues within the PTB domain binding site (facing away) and putative PH domain binding site are colored dark gray and numbered; residues at the interface between the two domains are colored according to percent buried: red, %; orange, 25-50%; and yellow, 1-25%. (B) The PH/PTB domain interface viewed as an open book. The PH and PTB domains each were rotated 90o, relative to their orientations in A, but in opposite directions to expose the buried surface between them. Elements of secondary structure and contact residues are labeled. [PNAS 96, , 1999]")
34
WW domén Foszfo-szerin, foszfo-treonin kötő modul
WW domains are compact 38 amino acid residue units that fold into a three-stranded b-sheet structure. A flat binding surface for the Pro-rich ligand is formed by conserved hydrophobic residues. The domain name is derived from two conserved Trp residues spaced 20 to 22 residues apart within the consensus sequence. The figure shows the Pin1 WW domain. Domain Binding and Function WW domains are small 38 to 40 amino acid residue modules that have been implicated in binding to Pro-rich sequences. WW domains and SH3 domains can potentially bind overlapping sites. In addition, the Pin1 WW domain functions as a phosphoserine- or phosphothreonine-binding module, suggesting that certain WW domains have evolved an alternate mode of action. WW domains bind peptide ligands with dissociation constants in the mmolar range. 38-40 aminosav, b lemez, két konzervált triptofán (W-W) aminosav távolságra, Kd = mM, pSer, PxxP kötés WW domains are compact 38 amino acid residue units that fold into a three-stranded b-sheet structure. A flat binding surface for the Pro-rich ligand is formed by conserved hydrophobic residues. The domain name is derived from two conserved Trp residues spaced 20 to 22 residues apart within the consensus sequence. The figure shows the Pin1 WW domain. Domain Binding and Function WW domains are small 38 to 40 amino acid residue modules that have been implicated in binding to Pro-rich sequences. WW domains and SH3 domains can potentially bind overlapping sites. In addition, the Pin1 WW domain functions as a phosphoserine- or phosphothreonine-binding module, suggesting that certain WW domains have evolved an alternate mode of action. WW domains bind peptide ligands with dissociation constants in the mmolar range.
 aminosav távolságra, Kd = mM, pSer, PxxP kötés. WW domains are compact 38 amino acid residue units that fold into a three-stranded b-sheet structure. A flat binding surface for the Pro-rich ligand is formed by conserved hydrophobic residues. The domain name is derived from two conserved Trp residues spaced 20 to 22 residues apart within the consensus sequence. The figure shows the Pin1 WW domain. Domain Binding and Function. WW domains are small 38 to 40 amino acid residue modules that have been implicated in binding to Pro-rich sequences. WW domains and SH3 domains can potentially bind overlapping sites. In addition, the Pin1 WW domain functions as a phosphoserine- or phosphothreonine-binding module, suggesting that certain WW domains have evolved an alternate mode of action. WW domains bind peptide ligands with dissociation constants in the mmolar range.")
35
Szemikonzervatív, 38-40 aminosavból álló domén, WW konzervetív
WW domain [EMBO-J. 15(5) 1996] [Nature 382, , 1996] Szemikonzervatív, aminosavból álló domén, WW konzervetív Kompakt globuláris szerkezet hidrofób zsebbel, ez prolinban gazdag peptideket is köthet (xPPxY), néha a ligandum identikus az SH3 ligandumokkal (PxxP) Kompetíció SH3 doménnel. WW domének SH3-hoz hasonló funkció következtében szabályozhatják az előbbiek célfehérjéivel való kölcsönhatásást WW Domének, mint P-Ser- vagy P-Thr-kötő modulok: A WW domén (Proline-isomerase Pin1) és az ubiqutin ligáz egyik WW doménje P-Ser-Pro and P-Thr-Pro motívumot tartalmazó fehérjékhez kötődik Science 283(5406): , 1999
 1996] [Nature 382, , 1996] Szemikonzervatív, aminosavból álló domén, WW konzervetív. Kompakt globuláris szerkezet hidrofób zsebbel, ez. prolinban gazdag peptideket is köthet (xPPxY), néha a ligandum identikus az SH3 ligandumokkal (PxxP) Kompetíció SH3 doménnel. WW domének SH3-hoz hasonló funkció következtében szabályozhatják az előbbiek célfehérjéivel való kölcsönhatásást. WW Domének, mint P-Ser- vagy P-Thr-kötő modulok: A WW domén (Proline-isomerase Pin1) és az ubiqutin ligáz egyik WW doménje P-Ser-Pro and P-Thr-Pro motívumot tartalmazó fehérjékhez kötődik. Science 283(5406): ,")
36
Kilenc anti-paralel a helix, dimer, pSer peptideket köt
domén - töltés Kilenc anti-paralel a helix, dimer, pSer peptideket köt Magcitoplazma export Crystal structures of the mammalian tau and epsilon isoforms of the module show that each module is composed of nine anti-parallel alpha helices. Dimerization results in the formation of a negatively charged channel between the two monomers. This channel is lined with invariant residues that mediate interactions with a phosphoserine peptide target sequence. The figure shows the x dimer complexed with phosphoserine peptide containing the binding motif. proteins are 30 kDa polypeptides with nine closely related members in mammals. They are also found in plants and fungi. They are involved in regulating various pathways including signal transduction, apoptosis and passage through the cell cycle proteins form homo- and hetero-dimeric cup-like structures that bind to discrete phosphoserine-containing motifs. In some instances, proteins appear to export their binding partners from the nucleus to the cytoplasm in a phosphorylation- and Crm1-dependent manner. Binding partners: Function : Cdc25 tyrosine phosphatase Cell cycle regulation BAD (Bcl-XL binding partner) Regulation of apoptosis c-Raf Ser/Thr Kinase Regulation of kinase activity; PKC Ser/Thr Kinase Signal transduction MEKK1,2,3 Ser/Thr Kinase Signal transduction
 Regulation of apoptosis. c-Raf Ser/Thr Kinase Regulation of kinase activity; PKC Ser/Thr Kinase Signal transduction. MEKK1,2,3 Ser/Thr Kinase Signal transduction.")
37
BH domén GQAGRQLAIIGDDINR
The structure of Bcl-XL consist of two central a-helices (a5 (BH1) and a6), which are flanked on one side by a3 and a4 and on the other by a1 (BH4), a2 (BH3) and a7 (BH2). The two central helices contain predominantly hydrophobic residues, and are arranged in an antiparallell fashion. The figure shows the BH1 (yellow), BH2 (magenta), BH3 (orange) and BH4 (cyan) domains of Bcl-XL. The Bak peptide (red), GQAGRQLAIIGDDINR, interacts predominantly with residues in the BH2 and BH3 regions. Domain Binding and Function Bcl-2 homology (BH1-4) domains are found in proteins that inhibit apoptosis such as Bcl-2, Bcl-XL and Bcl-XW. Bcl-2 family members form homodimers and heterodimers between pro and anti apoptotic family members. Homodimerization of Bcl-2 involves a head- to- tail interaction. The N-terminal region, where the BH4 domain resides, interacts with the more distal region of Bcl-2 where BH1, BH2 and BH3 are located. The BH3 domain is required for dimerization and apoptosis induction. Conversely, Bcl-2/Bax heterodimerization involves a tail- to- tail interaction that requires the BH1, BH2, and BH3 region of Bcl-2 and a central region in Bax where the BH3 domain is located Bcl-XL Két antiparalel a helix: (BH1) centrális, hidrofób, + csatlakozó BH4, BH3, BH2 helixek (Bcl-XL) Anti-apoptotikus és pro-apoptotikus molekula család, homo és heterodimereket képez – apoptózis gátlás
 and a6), which are flanked on one side by a3 and a4 and on the other by a1 (BH4), a2 (BH3) and a7 (BH2). The two central helices contain predominantly hydrophobic residues, and are arranged in an antiparallell fashion. The figure shows the BH1 (yellow), BH2 (magenta), BH3 (orange) and BH4 (cyan) domains of Bcl-XL. The Bak peptide (red), GQAGRQLAIIGDDINR, interacts predominantly with residues in the BH2 and BH3 regions. Domain Binding and Function. Bcl-2 homology (BH1-4) domains are found in proteins that inhibit apoptosis such as Bcl-2, Bcl-XL and Bcl-XW. Bcl-2 family members form homodimers and heterodimers between pro and anti apoptotic family members. Homodimerization of Bcl-2 involves a head- to- tail interaction. The N-terminal region, where the BH4 domain resides, interacts with the more distal region of Bcl-2 where BH1, BH2 and BH3 are located. The BH3 domain is required for dimerization and apoptosis induction. Conversely, Bcl-2/Bax heterodimerization involves a tail- to- tail interaction that requires the BH1, BH2, and BH3 region of Bcl-2 and a central region in Bax where the BH3 domain is located. Bcl-XL. Két antiparalel a helix: (BH1) centrális, hidrofób, + csatlakozó BH4, BH3, BH2 helixek (Bcl-XL) Anti-apoptotikus és pro-apoptotikus molekula család, homo és heterodimereket képez – apoptózis gátlás.")
38
Receptor tirozin kinázok családjai
Six subfamilies of receptor tyrosine kinases. Only one or two members of each subfamily are indicated. Note that the tyrosine kinase domain is interrupted by a "kinase insert region" in some of the subfamilies. The functional significance of the cysteine-rich and immunoglobulinlike domains is unknown. Tirozin kinázokhoz kapcsolt receptorok: TCR, BCR, FcR, citokin receptorok

39
Emberi növekedési hormon kötődése receptorához:
receptor dimerizáció –aktiváció előfeltétele Receptor homodimer The three-dimensional structure of human growth hormone bound to its receptor. The hormone ( red) has cross-linked two identical receptors (one shown in green and the other in blue) to form a receptor homodimer. (It was entirely unexpected that a monomeric ligand such as growth hormone would cross-link its receptors, as it requires that the two identical receptors recognize different parts of the hormone.) Ligand-induced dimerization is thought to bring together the cytoplasmic domains of the two single-pass, transmembrane receptor proteins. This in turn activates a nonreceptor tyrosine kinase (not shown). The structures shown were determined by x-ray crystallographic studies of complexes formed between the hormone and the extracellular receptor domain produced by recombinant DNA technology.
 has cross-linked two identical receptors (one shown in green and the other in blue) to form a receptor homodimer. (It was entirely unexpected that a monomeric ligand such as growth hormone would cross-link its receptors, as it requires that the two identical receptors recognize different parts of the hormone.) Ligand-induced dimerization is thought to bring together the cytoplasmic domains of the two single-pass, transmembrane receptor proteins. This in turn activates a nonreceptor tyrosine kinase (not shown). The structures shown were determined by x-ray crystallographic studies of complexes formed between the hormone and the extracellular receptor domain produced by recombinant DNA technology.")
40
Kináz funkciójának vizsgálata
A jelátadás gátlása mutáns receptor felesleggel: domináns negatív gátlás Figure Inhibition of signaling through normal receptor tyrosine kinases by an excess of mutant receptors. (A) In this example, the normal receptors dimerize in response to ligand binding. The two kinase domains cross-phosphorylate each other, increasing the activity of the kinase domains, which can now further activate the receptor dimer by phosphorylating other sites on the receptors. (B) The mutant receptor with an inactivated kinase domain can dimerize normally, but it cannot cross-phosphorylate a normal receptor in a dimer. For this reason, the mutant receptors, if present in excess, will block signaling by the normal receptors—a process called dominant-negative inhibition. Cell biologists frequently use this strategy to inhibit a specific type of receptor tyrosine kinase in a cell to determine its normal function
 In this example, the normal receptors dimerize in response to ligand binding. The two kinase domains cross-phosphorylate each other, increasing the activity of the kinase domains, which can now further activate the receptor dimer by phosphorylating other sites on the receptors. (B) The mutant receptor with an inactivated kinase domain can dimerize normally, but it cannot cross-phosphorylate a normal receptor in a dimer. For this reason, the mutant receptors, if present in excess, will block signaling by the normal receptors—a process called dominant-negative inhibition. Cell biologists frequently use this strategy to inhibit a specific type of receptor tyrosine kinase in a cell to determine its normal function.")
41
RTK aktiválás bivalens ellenanyagokkal

42
Receptor tirozin kinázok (RTK) közvetítenek:
-sejtek növekedését, túlélését -differenciálódást -sejt mozgását -metabolizmusát befolyásoló jeleket ==> Szabályozás hiányában a RTK fokozott aktivitása malignus transzformációhoz vezet ==> Csökkent RTK aktivitás a sejtek szaporodásának és fejlődésének hibájához és metabolizmus zavarán alapuló, súlyos betegségekhez vezethet. -A receptorok 14 családba sorolhatók, az azonos család receptorai között ligandumaikat tekintve hasonlóság van

43
Hogyan történik a RTK jeltovábbítása?
A receptor tirozin kinázokon kialakuló szignaloszóma segítségével:

44
RTK közvetítet jelátvitel kezdeti lépései
A receptor tirozin kinázok specifikus felismerést biztosító kötő- és gyüjtőhelyei a jeltovábbító molekuláknak

45
Az intracelluláris régió szub-doménei: Membránközeli régió:
Bázikus maradékok, melyek kölcsönhatásba lépnek a negatív töltésű foszfolipidekkel: „stop transfer” Internalizációs jel Ser /Thr maradékok: foszforilálódhatnak, és szabályozhatják a RTK aktivitását Tirozin kináz domén: ATP-kötő hely autofoszforiláció "activation loop„ „Kináz insert” régió, sok tirozint tartalmaz, ezek foszforilációja kötőhelyeket biztosít az effektor fehérjék számára Szubsztrát kötő régió: "catalytic loop” C terminális régió : számos tirozint tartalmaz Foszforilációt követően dokkoló helyül szolgálnak az effektor fehérjék számára. plazmamembrán

46
Az intracelluláris receptor régió
Tirozin kináz domént tartalmaz (ez különbözteti meg a RTK család tagjait más receptoroktól) Az intracelluláris régió szub-doménekre oszlik : i. Kb. 50 aminosav – membránközeli régió: Bázikus maradékokat tartalmaz, melyek kölcsönhatásba lépnek a negatív töltésű foszfolipidekkel és „stop transfer” szignált hordoz Internalizációs jelet tartalmaz ( NPXY motívum, LDL receptor GPLY motívum) Ser /Thr maradékok, amelyek foszforilálódhatnak, és szabályozhatják a RTK aktivitását
 Az intracelluláris régió szub-doménekre oszlik : i. Kb. 50 aminosav – membránközeli régió: Bázikus maradékokat tartalmaz, melyek kölcsönhatásba lépnek a negatív töltésű foszfolipidekkel és „stop transfer szignált hordoz. Internalizációs jelet tartalmaz ( NPXY motívum, LDL receptor GPLY motívum) Ser /Thr maradékok, amelyek foszforilálódhatnak, és szabályozhatják a RTK aktivitását.")
47
ii. Tirozin kináz domén ATP-kötő hely Tyr autofoszforilációs hely "activation loop" Szubsztrát kötő régió "catalytic loop” iii. „kináz insert” régió, A PDGF/FGF családban ez a régió sok tirozint tartalmaz Ezek foszforilációja kötőhelyeket biztosít az effektor fehérjék számára iv. C terminális régió : számos tirozint tartalmaz Foszforilációt követően dokkoló helyül szolgálnak az effektor fehérjék számára.

48
Foszforilált tirozin maradékok biztosítják a jeltovábbításban
Vad típus Mutált PDGFR Kináz aktív Ca2+, IP3 válasz Sejtszaporodás: MIÉRT?: PI3-K Q: What is the Role of Autophosphorylation that occurs subsequent to dimer formation? i. To activate the RTK (e. g. HGF; insulin, FGF, IGF-1 receptors, with increase in Vmax with no change in Km) ii. Generate docking sites for downstream effector molecules. [JBC, 258, , 1983] [PNAS, 80, , 1983] [MCB, 16, , 1996] Q: What is the mechanism of autophosphorylation ? intramolecular "trans" mechanism (IR, IGF-1R) in which one receptor-half initially phosphorylates the other. [JBC, 267, , 1992] Intermolecular mechanism (EGFR, FGFR) [Cell 86, , 1996] Foszforilált tirozin maradékok biztosítják a jeltovábbításban résztvevő fehérjék kötődését a receptorhoz, amely a teljes biológiai válasz előfeltétele
 ii. Generate docking sites for downstream effector molecules. [JBC, 258, , 1983] [PNAS, 80, , 1983] [MCB, 16, , 1996] Q: What is the mechanism of autophosphorylation intramolecular trans mechanism (IR, IGF-1R) in which one receptor-half initially phosphorylates the other. [JBC, 267, , 1992] Intermolecular mechanism (EGFR, FGFR) [Cell 86, , 1996] Foszforilált tirozin maradékok biztosítják a jeltovábbításban. résztvevő fehérjék kötődését a receptorhoz, amely a teljes. biológiai válasz előfeltétele.")
49
Five parallel intracellular signaling pathways activated by G-protein-linked receptors, receptor tyrosine kinases, or both Figure Five parallel intracellular signaling pathways activated by G-protein-linked receptors, receptor tyrosine kinases, or both. In this schematic example, the five kinases (shaded yellow) at the end of each pathway phosphorylate target proteins (shaded red), some of which are phosphorylated by more than one of the kinases. The specific phospholipase C activated by the two types of receptors is different: G-protein-linked receptors activate PLC-β, whereas receptor tyrosine kinases activate PLC-γ (not shown).
 at the end of each pathway phosphorylate target proteins (shaded red), some of which are phosphorylated by more than one of the kinases. The specific phospholipase C activated by the two types of receptors is different: G-protein-linked receptors activate PLC-β, whereas receptor tyrosine kinases activate PLC-γ (not shown).")
Hasonló előadás















>")
>")

 kialakulása Genetikai, Sejt- és Immunbiológiai Intézet Falus András.>")
