Előadást letölteni
Az előadás letöltése folymat van. Kérjük, várjon

1
Sejtmembrán szerkezete és műküdése
Dr. habil. Kőhidai László egyetemi docens SE, Genetikai, Sejt- és Immunbiológiai Intézet 2008.

2
Prokaryota sejt Eukaryota sejt szekréciós szemcse színtest riboszóma
citoplazma pílusok riboszóma sejtmembrán sejtfal tok ostor Eukaryota sejt szekréciós szemcse színtest riboszóma lizoszóma pinocitotikus vezikulum csillók mikro- bolyhok centriolumok Golgi apparátus sima felszínű endoplazmatikus hálózat sejtmagvacska kromatin állomány mitochondrium nukleoplazma citoplazma sejtmag sejtmag- membrán pórusok durva felszínű thylakoid sejtmembrán mikrotubulus filamentum

3
Prokaryota sejt Eukaryota sejt szekréciós szemcse színtest riboszóma
citoplazma pílusok riboszóma sejtmembrán sejtfal tok ostor Eukaryota sejt szekréciós szemcse színtest riboszóma lizoszóma pinocitotikus vezikulum csillók mikro- bolyhok centriolumok Golgi apparátus sima felszínű endoplazmatikus hálózat sejtmagvacska kromatin állomány mitochondrium nukleoplazma citoplazma sejtmag sejtmag- membrán pórusok durva felszínű thylakoid sejtmembrán mikrotubulus filamentum

4
Kompartmentalizálódás
Az egyes biokémiai folyamatok membránokkal határolt terekbe történő szétválasztása – optimális környezeti feltételek biztosítása

5
A citoplazmatikus membránok fő funkciói
Határhártya képzése – tápanyagok be és kifelé irányuló tarnszportját segíti elő Fehérjék kihorgonyzása – a traszport, bioenergetikai folyamatok, szignálok észlelése és kemotaxis Figure: 04-21 Caption: The major functions of the cytoplasmic membrane. Energia raktározás - proton gradiens generálása

6
A membránok funkciói 1. Határhártyát képeznek és szelektív átjárhatóságú felszínek (ld.permeabilitás) – sejt membrán/plazmamembrán és intracellularis membránok – sejt organellumok 2. Specifikus funkciókat töltenek be – a kapcsolódó proteinek alapján megkülönböztethetők (integráns vagy perifériás fehérjék). Glucose phosphatase – az ER-membrán-kötött enzym és egyben annak markere is. 3. A membránok szabályozzák egyes anyagok transzportját – a. Egyszerű diffúzió (magas cc.->alacsony cc.): H2O, O2, CO2, ethanol b. Facilitált diffúzió (magas cc.->alacsony cc.) transzport proteinek segítségével: cukrok, aminosavak c. “Pumpa mechanizmusok” proteinjei, melyek az ion-gradienseket tartják fenn: alacsony cc.-> magas cc. Cukrok, aminosavak sejtbe juttatása koncentráció gradiens ellenében is. d. Endocytosis és exocytosis, stb
 – sejt membrán/plazmamembrán és intracellularis membránok – sejt organellumok. 2. Specifikus funkciókat töltenek be – a kapcsolódó proteinek alapján megkülönböztethetők (integráns vagy perifériás fehérjék). Glucose phosphatase – az ER-membrán-kötött enzym és egyben annak markere is. 3. A membránok szabályozzák egyes anyagok transzportját – a. Egyszerű diffúzió (magas cc.->alacsony cc.): H2O, O2, CO2, ethanol. b. Facilitált diffúzió (magas cc.->alacsony cc.) transzport proteinek segítségével: cukrok, aminosavak. c. Pumpa mechanizmusok proteinjei, melyek az ion-gradienseket tartják fenn: alacsony cc.-> magas cc. Cukrok, aminosavak sejtbe juttatása koncentráció gradiens ellenében is. d. Endocytosis és exocytosis, stb.")
7
A membránok funkciói 4. Elektromos és kémiai jelek detektálása és továbbítása – szignál transzdukció: külső jelek fogadása és továbbítása. Pl.: Ösztrogén – hidrofób karaktere miatt képes átjutni a membránon; más anyagok hatásához receptorhoz való kötődés szükséges, mely másodlagos hírvivőket generál. A jel a sejtmagi szintig is eljuthat és befolyásolhatja a génexpressziót. 5. Sejt-sejt kommunikáció közvetítése – gap junction (állati sejtek) és plasmodesmata (növényi sejtek) intercelluláris kommunikációt tesznek lehetővé.
 és plasmodesmata (növényi sejtek) intercelluláris kommunikációt tesznek lehetővé.")
8
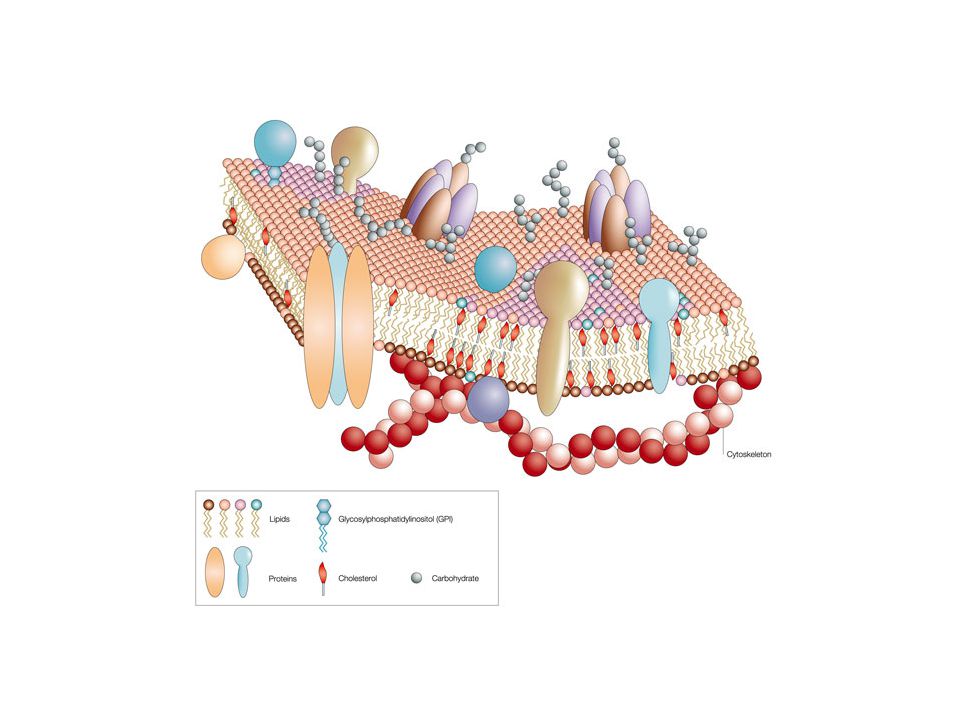
Membrán-modellek Lipid-oldékony anyagok gyorsan bejutnak a sejtbe
Benzin-lipid keverék, a benzin elpárolgása után molekuláris lipid film keletkezik Benzin-oldékony lipidek két rétegből álló filmet alkotnak víz felszínén A fehérjék integráns részei a sejt membránnak. A lipid bilayert kívül-belül proteinek rétege fedi. Részben magyarázza a proteinek, cukrok, ionok és más hidrofil anyagok gyors átjutását. Elektronmikroszkóp felfedezése. A sejtek plazmamembránnal burkoltak. „Unit-membrane”modell. Proteinek mozaikos elrendeződése a membránban. Egyes proteinek átérik a membránt, transzmembrán proteinek

9
Fluid mozaik membrán modell
Singer – Nicolson 1972 vvt. membrán

10
Membrán modellek – Folyékony mozaik modell:
A proteinek mozaikosan, elszórtan és beágyazottan helyezkednek el a folyékony lipid kettősrétegben Bacteriorhodopsin: az első transzmembrán doménnel leírt protein, mely retinol-szerű részével energia megkötő képességű is egyben. A napfény energiája megkötődés után szerkezeti változást idéz elő, mely protonok sejtből tőrténő kijuttatását eredményezi. A kialakuló proton-gradiens energia-forrásként szolgál. A legtöbb membrán felépítésében lipidek, fehérjék és szénhidrátok is részt vesznek.

11
Foszfolipidek általános szerkezeti felépítése

12
A membránt felépítő fő lipid-komponensek
Foszfolipidek Glikolipidek Egyéb membrán-lipidek (nem foszfolipidek) Cholesterin Cerebrozid Szfingolipidek Ceramid
 Cholesterin. Cerebrozid. Szfingolipidek. Ceramid.")
13
Membrán lipidek: a modell „fluiditásáért” felelős elemek
A membránok alkotásában több féle lipid vesz részt Foszfolipidek Foszfogliceridek Szfingolipidek Különböző membránok foszfolipid összetétele

14
Glikolipidek Cerebrozidok és gangliozidok Tay-Sachs betegség: β–N-acetilhexoaminidáz hiányos lizoszómák, gangliozidok degradációja károsodik; gangliozid akkumuláció az idegrendszerben. Szteroidok (nem található a prokaryotákban vagy a mitochondrium, színtest belső membránjaiban) Cholesterin Fitoszterin A baktériumok és a cyanobaktériumok membránjaiban szteroid-szerű molekulák találhatók – haponoid (petroleum depozitokban gazdag; korai prokaryoták membrán componensei?)
 Cholesterin. Fitoszterin. A baktériumok és a cyanobaktériumok membránjaiban szteroid-szerű molekulák találhatók – haponoid (petroleum depozitokban gazdag; korai prokaryoták membrán componensei )")
15
Vékonyréteg-kromatográfia – a lipid analízis fontos eszköze:
Álló fázis – vékony szilikagél, hidrofil rétege üveg vagy fém lemezen Mozgó fázis – oldószer chloroform, methanol, és water keveréke Cholesterin Phosphatidylethonolamine (PE) Phosphatidylcholine (PC) Phosphatidylserine (PS) Alacsony affinitás Magas affinitás Zsírsavak a membrán szerkezete és működése szempontjából is jelentősek A zsírsavak minden membrán-lipid alkotói kivéve a szteroidokat. A hidrofób szénhidrogének általában 16- vagy 18-C hosszúak és a lipid kettősréteg stabilitását adják. Telített zsírsavak: palmitinsav(16C) és sztearinsav (18C) Telítetlen zsírsavak: Olajsav és linolsav (mk. 18C)
 Phosphatidylcholine (PC) Phosphatidylserine (PS) Alacsony affinitás. Magas affinitás. Zsírsavak a membrán szerkezete és működése szempontjából is jelentősek. A zsírsavak minden membrán-lipid alkotói kivéve a szteroidokat. A hidrofób szénhidrogének általában 16- vagy 18-C hosszúak és a lipid kettősréteg stabilitását adják. Telített zsírsavak: palmitinsav(16C) és sztearinsav (18C) Telítetlen zsírsavak: Olajsav és linolsav (mk. 18C)")
16
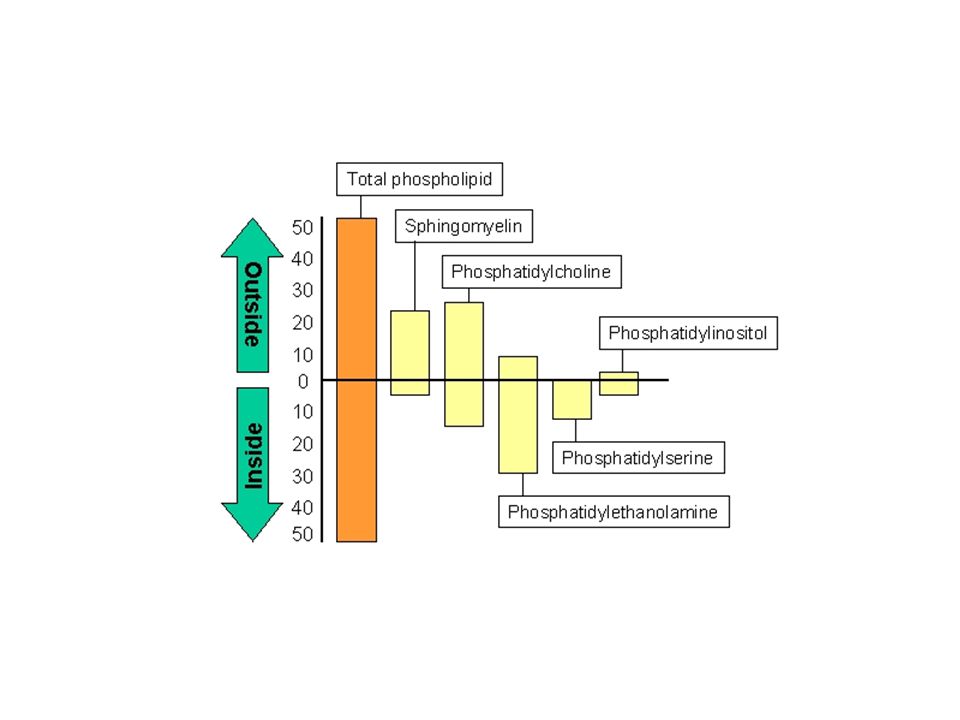
Membrán- aszimmetria: a lipidek döntő hányada egyenlőtlen arányban oszlik meg a két lipid-monolayerben - mennyiségi és minőségi eltérések Az aszimmetria a lipidek típusainak eltérőségében és a telítettség fokának különbözőségében testesül meg. Az aszimmetria a membránok biogenezise során alakul ki. A kialakult aszimmetria termodinamikai okoból gátolja a lipidek cseréjét a két felszín között. Külső membrán: glikolipidek – felismerő folyamatok és jeltovábbítás Belső membrán: a sejt belső terébe történő jeltovábbításban fontosak -foszfatidiletanolamin, foszfatidilinozitol, foszfatidilszerin Lipidek mozgása a membránban: “flip-flop”, vagy transzverz diffúzió rotáció laterális diffúzió A flip-flop mozgás ritka, de a biológiai membránokban az ú.n. foszfolipid transzlokátorok vagy flippázok segítik elő. Ezért gyakoribb a foszfolipid transzlokáció a biológiai membránokban, mint a mesterséges lipid bilayerekben.

17
A membrán aszimmetria kialakulása és fenntartása:
Foszfolipidek szintézise az ER membránhoz kötötten zajlik. A foszfolipidek az ER citoplazmatikus felszínéhez kapcsolódnak A flippáz juttatja át az egyes lipideket a bilayer másik oldalára.

18
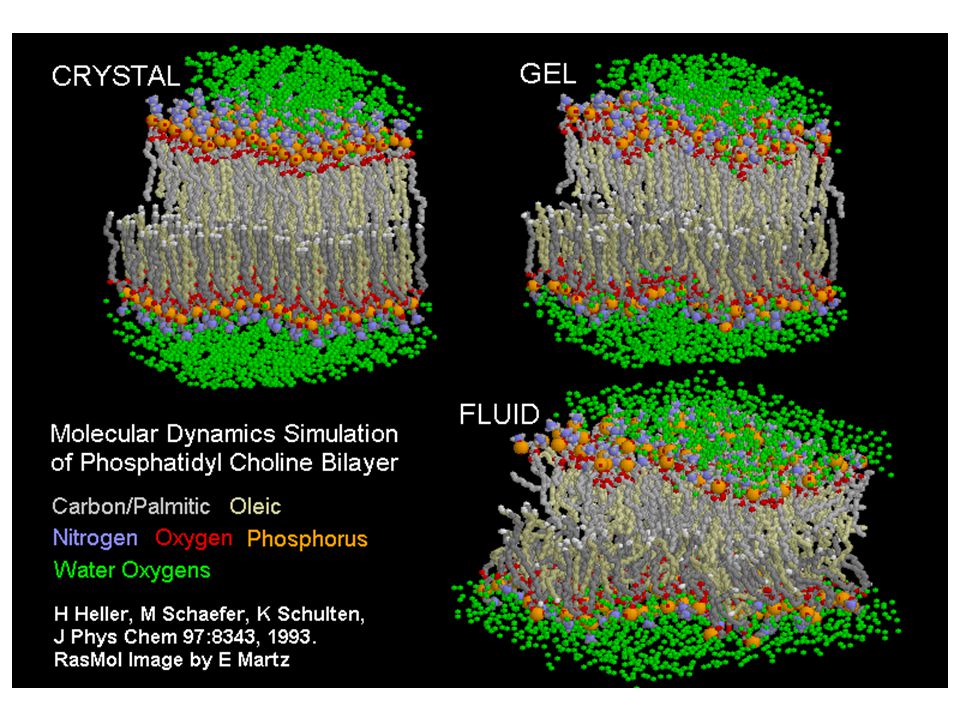
A lipid bilayer fluiditásának vizsgálata
Laterális diffúzióval a lipidek és proteinek pl.: egy 800 M.S.-jú lipid néhány mM (egy baktérium hosszával azonos távolságot) tud elmozdulni 1 mp alatt! A proteinek nagy méretük miatt lassabban mozognak. Vizsgálati eljárás: fluorescence recovery after photobleaching (FRAP) technika A membrán megfelelően csak „fluid” állapotában működik Fázis-átmeneti hőmérséklet (Tm): Fázis-átmenet: gél-szerű állapot -> fluid „Differential scanning calorimetry” Calometer: a hőabszorpció mérésére szolgáló kamra. A görbe csúcsának helye jelzi a Tm-et. Telítetlen: alacsony Tm Telített: magas Tm
 tud elmozdulni 1 mp alatt! A proteinek nagy méretük miatt lassabban mozognak. Vizsgálati eljárás: fluorescence recovery after photobleaching (FRAP) technika. A membrán megfelelően csak „fluid állapotában működik. Fázis-átmeneti hőmérséklet (Tm): Fázis-átmenet: gél-szerű állapot -> fluid. „Differential scanning calorimetry Calometer: a hőabszorpció mérésére szolgáló kamra. A görbe csúcsának helye jelzi a Tm-et. Telítetlen: alacsony Tm. Telített: magas Tm.")
19
Membrán-fluiditás és zsírsavösszetétel kapcsolata
Két fő szempont: lánchossz és telítettség mértéke 18-C lipidek 70oC -11oC 76oC 32oC Membrán-lipidek gyakran 1 telített és 1 telítettlen zsírsavból épülnek fel Szteroidok hatása a membránfluiditásra Szteroidok – cholesterin és phytosterin. A szteroidok a teljes membrán-lipid tartalom 50%-át is kitehetik Cholesterin – hidrogén kötéseket alakít ki a szomszéd foszfolipidek oxigén atomjaival. A cholesterin paradox hatása: magas hőmérsékleten csökkenti a membrán fluiditást (a rigid gyűrűk hatása) míg alacsony hőmérsékleten növeli a fluiditást (a szomszédos szénhidrogén láncok természetes elrendeződését gátolva) AZONBAN, a szteroidok csökkentik a lipid bilayer permeabilitását – a foszfolipidek közötti teret kitöltve. Az élőlények szabályozni képesek a membrán fluiditást Poikelotherm-ek (baktérium, gomba, növény, kétéltűek stb.) vs. homeotherm-ek (melegvérű állatok) Homeoviszkózus adaptáció – a membrán viszkozitása megtartott a változó hőmérséklet ellenére – terminális C-k eltávolításával zsírsavláncok rövidítése, ill. telítetlenné tétele (pl.: E. coli desaturáz).
 míg alacsony hőmérsékleten növeli a fluiditást (a szomszédos szénhidrogén láncok természetes elrendeződését gátolva) AZONBAN, a szteroidok csökkentik a lipid bilayer permeabilitását – a foszfolipidek közötti teret kitöltve. Az élőlények szabályozni képesek a membrán fluiditást. Poikelotherm-ek (baktérium, gomba, növény, kétéltűek stb.) vs. homeotherm-ek (melegvérű állatok) Homeoviszkózus adaptáció – a membrán viszkozitása megtartott a változó hőmérséklet ellenére – terminális C-k eltávolításával zsírsavláncok rövidítése, ill. telítetlenné tétele (pl.: E. coli desaturáz).")
20
A membrán fluiditásának befolyásolása:
Foszfolipidek: lánchossz telítettség Cholesterin-tartalom

21
Lipid raft-ok: - cholesterinben gazdag - mikro-domainek
glikolipidek+szfingolipidek a raftok jelen vannak a sejt felszíni membránján és a Golgiról lefűződő membránokon jelentőségük: sejtmembrán egyes rétegei közötti szabadenergia különbség csökkentése membránhoz kötött transzport folyamatok Más nevei a szakirodalomban: detergent-insoluble glycolipid-enriched complexes (GEM vagy DIG) detergent resistant membranes (DRMs). Kimutatás: fluoreszcens mikroszkópia – GM1 gangliozidok; AFM
 detergent resistant membranes (DRMs). Kimutatás: fluoreszcens mikroszkópia – GM1 gangliozidok; AFM.")
22
E felszín (exoplazmatikus oldal) és P felszín (protoplazmatikus oldal)
Membrán-proteinek: a modell “mozaik”-jellege Freezing fracturing: gyors fagyasztás (folyékony nitrogén) után gyémánt-késsel történő feltárás E felszín (exoplazmatikus oldal) és P felszín (protoplazmatikus oldal)
 után gyémánt-késsel történő feltárás. E felszín (exoplazmatikus oldal) és P felszín (protoplazmatikus oldal)")
23
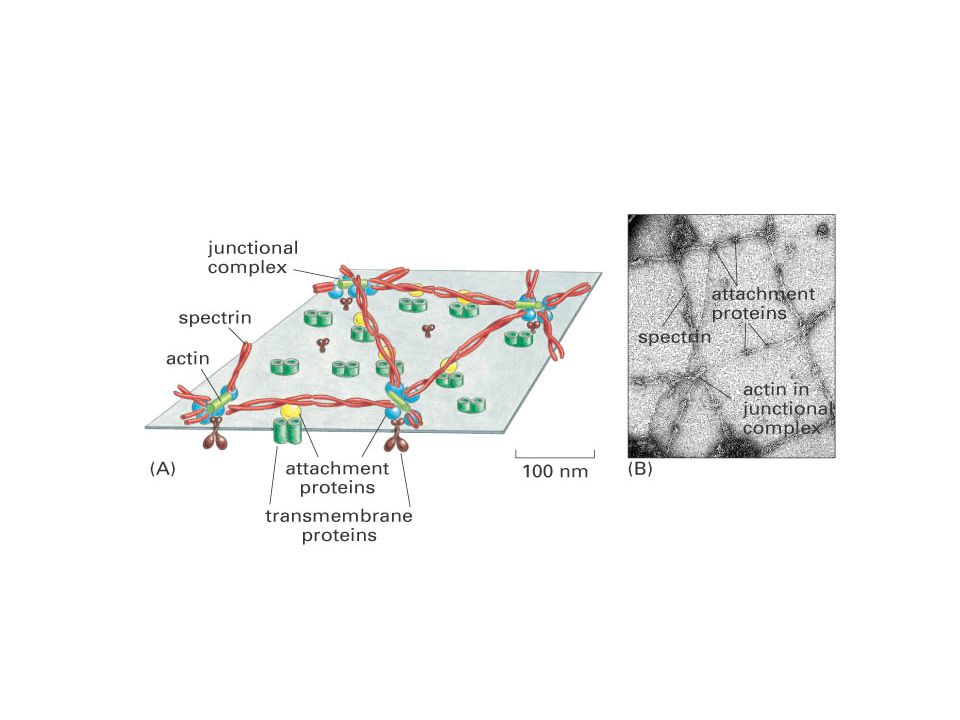
Membrán proteinjei: - integráns - perifériális
Integráns membrán proteinek: ~20 aminosavból álló, transzmembrán szegmentum. Nehezen izolálható detergens kezelés kell az extrakciójához.. Intergráns monotóp proteinek Transzmembrán proteinek: singlepass multipass multi-subunit Membrán proteinjei: - integráns - perifériális - lipidek által kihorgonyzott Glycophorin (vvt.) Hydropathy plot Bacteriorhodopsin
 Hydropathy plot. Bacteriorhodopsin.")
24
Lipidekhez kötött membrán proteinek: kovalens kötés a lipiddel.
Perifériásl membrán proteinek a membrán felszínekhez gyenge elektrostatikus erők vagy hidrogén kötések rögzítik. Kapcsolat az integráns proteinek hidrofil részeivel vagy a membrán lipidek poláris feji részeivel. pl.: spectrin és ankyrin a vvt. plazmamembránjában. Lipidekhez kötött membrán proteinek: kovalens kötés a lipiddel. Zsírsav vagy prenyl mint kihorgonyzó GPI-kapcsolófdás (glycosylphosphatidyinositol, GPI) A proteineket SDS-polyacrylamid gél elektroforézissel választhatjuk el Membrán proteinek izolálása Perifériás membrán proteinek: a pH vagy az ionos kötések megváltoztatása, calcium eltávolítása –chelatiós-agensek (kationos-kötés), vagy urea (a hidrogén kötéseket bontja). A lipid-kötött proteinek hasonló módon izolálhatók, mint a perifériás proteinek, mivel itt is kovalens kötéseket kell bontani. Az integráns membránproteinek kivonása nehezebb, detergens igényes feladat. A detergens a hidrofób kötéseket bontja és a lipid bilayerrel lép kölcsönhatásba ld. SDS (sodium dodecyl sulfate) kezelés SDS-polyacrylamid gél elektroforézis: Elektroforézis elektromos tér alkalmazása elektromosan töltött molekulák elválasztására. A vándorlási sebesség a töltéstől és a mérettől függ. SDS feltárás (proteinek a negatívan töltött SDS-sel fedettek) -> elektroforézis -> festés Coomassie brilliant blue-val.
 A proteineket SDS-polyacrylamid gél elektroforézissel választhatjuk el. Membrán proteinek izolálása. Perifériás membrán proteinek: a pH vagy az ionos kötések megváltoztatása, calcium eltávolítása –chelatiós-agensek (kationos-kötés), vagy urea (a hidrogén kötéseket bontja). A lipid-kötött proteinek hasonló módon izolálhatók, mint a perifériás proteinek, mivel itt is kovalens kötéseket kell bontani. Az integráns membránproteinek kivonása nehezebb, detergens igényes feladat. A detergens a hidrofób kötéseket bontja és a lipid bilayerrel lép kölcsönhatásba ld. SDS (sodium dodecyl sulfate) kezelés. SDS-polyacrylamid gél elektroforézis: Elektroforézis elektromos tér alkalmazása elektromosan töltött molekulák elválasztására. A vándorlási sebesség a töltéstől és a mérettől függ. SDS feltárás (proteinek a negatívan töltött SDS-sel fedettek) -> elektroforézis -> festés Coomassie brilliant blue-val.")
25
Membrán-proteinek izolálása és vizsgálata:
Az alkalmazott detergensek a proteinek körüli foszfolipid szerkezetet utánozzák hidrifób hidrofil

26
??? A membrán-proteinek számos funkciót tölthetnek be
Enzim: Pl.: glukóz foszfatáz (az ER-mal asszociált) és glicerinaldehid-3- foszfát dehidrogenáz (GPD), a vvt. Perifériás proteinjei a vér glukóz-metabolizmusában vesznek részt. Transzport proteinek pl.: transzport ATPázok ATP energiáját felhasználva juttatnak át ionokat a membránon. Receptorok: pl.: hormonok és növekedési faktorok Egyéb: endocitóziss és exocitózis; targeting, szortírozás, fehérjék módosítása az ER vagy a Golgi-ban; fény érzékelése; a sejtmembrán szerkezetének stabilizálása és alak meghatározás (ankyrin, spectrin). TRANSZPORTER KIHORGONYZÁS RECEPTOROK ENZYMEK Transzmembrán (intergráns) Single-pass Multi-pass Membrán-asszociált (perifériás) Kovalensen kötött vs nem-kovalens ???
 és glicerinaldehid-3- foszfát dehidrogenáz (GPD), a vvt. Perifériás proteinjei a vér glukóz-metabolizmusában vesznek részt. Transzport proteinek pl.: transzport ATPázok ATP energiáját felhasználva juttatnak át ionokat a membránon. Receptorok: pl.: hormonok és növekedési faktorok. Egyéb: endocitóziss és exocitózis; targeting, szortírozás, fehérjék módosítása az ER vagy a Golgi-ban; fény érzékelése; a sejtmembrán szerkezetének stabilizálása és alak meghatározás (ankyrin, spectrin). TRANSZPORTER. KIHORGONYZÁS. RECEPTOROK ENZYMEK. Transzmembrán (intergráns) Single-pass Multi-pass. Membrán-asszociált (perifériás) Kovalensen kötött vs nem-kovalens.")
28
A membrán-proteinek bilayerbe ágyazódása
Az a-helikális szerkezet igen gyakori. Ennek oka, hogy az így expresszált hidrofób aminosavak felszínre kerülésével a proteinek jól rögzülnek a membrán hidrofób rétegében Az a-helikális szerkezetek hidrofil komponensei a pórusképzésben fontosak

29
A sejtmembrán szénhidrát komponensei
Funkciók: Felszíni védelem Felismerés Sejtadhézió - -- extracellularis matrix

30
A membrán-proteinek nagy része glikozilált
Emberi vvt. membrán: 52% proteinek, 40% lipidek és 8% szénhidrátok. A szénhidrátok glikolipid (ritka) és glikoprotein (gyakori) formában találhatók a membránban Glikoproteinek: N-kötöttek az Asp oldalláncának amino csoportján keresztül O-kötöttek – Ser vagy Thr hidroxil csoportjaihoz kötött szénhidrátok Egyes O-kötött szénhidrát hidroxi-Lys vagy hidroxi-Pro hidroxil csoportján keresztül kapcsolódik
 és glikoprotein (gyakori) formában találhatók a membránban. Glikoproteinek: N-kötöttek az Asp oldalláncának amino csoportján keresztül. O-kötöttek – Ser vagy Thr hidroxil csoportjaihoz kötött szénhidrátok. Egyes O-kötött szénhidrát hidroxi-Lys vagy hidroxi-Pro hidroxil csoportján keresztül kapcsolódik.")
31
A proteinekhez kapcsolódó szénhidrátok hossza: 2-60 egység, egyenes v
A proteinekhez kapcsolódó szénhidrátok hossza: 2-60 egység, egyenes v. lineáris Gyakori felépítő elemek: galaktóz, mannóz, N-acetilglukózamin és sziálsav A vvt. integráns proteinjének (glycophorin) szénhidrát láncainak (16 ilyen láncból 15 O-kötött és 1 N-kötött) terminális pozíciójában negatívan töltöt sziálsav található – ez csökkenti a vvt.-k összetapadási hajlamát és a vér viszkozitását. A glikoproteinek elsődleges szerepet töltenek be a sejtmembránok sejt-sejt felismerő funkciójában. Kísérletek: Lectinek: növényi (állati) fehérjék, melyek cukorspecifikus kötést alkotnak. Pl.: wheat germ agglytinin (búza) N-acetilglukózaminra végződő oligoszaccharidokhoz kötődik, míg a „jack bean”-ből kivont concanavalin az oligoszaccharidok belső mannóz csoportjait ismeri fel. Ferritin, egy vas-tartalmú fehérje, melyet glycoproteinekhez kötnek és elektrondenzitásuk révén a membránhoz való kötődés így jól detektálható.
 szénhidrát láncainak (16 ilyen láncból 15 O-kötött és 1 N-kötött) terminális pozíciójában negatívan töltöt sziálsav található – ez csökkenti a vvt.-k összetapadási hajlamát és a vér viszkozitását. A glikoproteinek elsődleges szerepet töltenek be a sejtmembránok sejt-sejt felismerő funkciójában. Kísérletek: Lectinek: növényi (állati) fehérjék, melyek cukorspecifikus kötést alkotnak. Pl.: wheat germ agglytinin (búza) N-acetilglukózaminra végződő oligoszaccharidokhoz kötődik, míg a „jack bean -ből kivont concanavalin az oligoszaccharidok belső mannóz csoportjait ismeri fel. Ferritin, egy vas-tartalmú fehérje, melyet glycoproteinekhez kötnek és elektrondenzitásuk révén a membránhoz való kötődés így jól detektálható.")
32
Membrán proteinek jelölése fluoreszcensen jelzett antitesttel Vírus
A membránproteinek mobilitása igen eltérő lehet Egyes proteinek szabadon elmozdulnak, míg mások kötöttek. A mobilitás kimutatására felhasznált technika: sejtfúzió Membrán proteinek jelölése fluoreszcensen jelzett antitesttel Vírus indukálta hibrid sejt fúzió A proteinek néhány perc múlva keveredni kezdenek 40 perc múlva a keveredés tejles Egér sejt Emberi sejt

33
A sejtmembrán proteinjeinek mozgása korlátozott:
Pl. bélhámsejtek vagy mirigyhám polarizált felszíne Receptorok, transzporterek, sejtkapcsoló struktúrák polarizált elhelyezkedése, megjelenése.

34
A membránok dinamikus jellege
bilayer 5 nm Számos sejtélettani folyamat során változik a membrán alakja. Az alkotó elemkre ható erők módosítják a membrán kiterjedését. A komponensekre ható feszítő és nyújtó-erők magyarázzák a mebrán alakváltozásait. A membránhoz kötött enzimek fenti hatásokat módosíthatják.

35
Konstans lipid denzitás
A membrán nyújtása A membrán hajlítása konstans terület/fej NINCS nyújtás RELAXÁLT ÁLLAPOT Konstans lipid denzitás Nyújtás - feszülés NEM-RELAXÁLT ÁLLAPOT Membrán aszimmetria d Lipid denzitás

36
(Aminophospholipid translocase)
A diffúzió és a hajlat kialakulás egyaránt az elasztikus stressz csökkentését eredményezi Flippáz (Aminophospholipid translocase) Nyújtás Kontrakció
 Nyújtás. Kontrakció.")
37
A denzitás és a hajlatok a deformált membránra nézve
A perturbáció foka Átfordulások száma Konzerválás A denzitás és a hajlatok egységnyiek a deformált membránra nézve Oldószer-áram Membrán-áram

38
A MEMBRÁN SPONTÁN HAJLATOKAT KÉPEZ
A membránba beépülő elemek pl. proteinek “spontán hajlatok”-at idéznek elő HA A RIGIDITÁS <0 A MEMBRÁN SPONTÁN HAJLATOKAT KÉPEZ

39
Összefoglalás

42
Hőmérséklet Szilárd Folyékony

43
Extracelluláris Intracelluláris

45
Források: Hardin, J., Kleinsmith, L., Becker, W.: World of the cell with free solutions, (Benjamin Cummings, 2002) Alberts, B., Johnson, A., Lew, J.: Essential Cell Biology, (Garland, 2004)
 Alberts, B., Johnson, A., Lew, J.: Essential Cell Biology, (Garland, 2004)")
Hasonló előadás







>")











.>")